Які водорості не мають джгутикових стадій. Жгутикові водорості. Ключові терміни та поняття
У всіх гетероконтів схоже влаштований джгутиковий апарат. Є 2 джгутики, причому на одному з них є дуже характерні трубчасті тричленові перисті вирости, або волоски - мастигонеми. Саме наявність двох нерівних джгутиків стало підставою для того, щоб назвати цю велику групу водних організмів. різножгутиковимиводоростями, або гетероконтами. Крім відмінностей у довжині, джгутики розрізняються і морфологічно: головний джгутик складається з осі та перисто розташованих на ній миготливих волосків, бічний джгутик гладкий бичевидний. Подібний також склад пігментів – хлорофіли аі зта численні каротиноїди, а також будова хлоропластів. Хлоропласти оточені чотирма мембранами, причому дві зовнішні мембрани пов'язані з ендоплазматичною мережею клітини.
У складі відділу Гетероконтні (Різножгутикові) водорості поєднують кілька груп водоростей у ранзі класів: Бурі, Золотисті, Жовтозелені., Рафідофітові, Синурові, Феотамнієві, Євстигмові, деякі раніше розглядалися як самостійні відділи.
Клас Бурі водорості– Phaeophyceae (Fucophyceae)
Назва класу «бурі водорості» відображає забарвлення їхнього талому (від грец. рhaios- Смаглявий). Забарвлення обумовлене наявністю великої кількості бурих та жовтих пігментів – крім хлорофілів аі зі β -каротину в їх хлоропластах є надлишок ксантофілів, особливо фукоксантину.
Багатоклітинні, переважно великі морські водорості, особливо поширені в холодних водах обох півкуль. Ні одноклітинні, ні колоніальні форми серед бурих водоростей невідомі. Відомо близько 1,8 тис. видів.
Будова клітини. Клітина у видів бурих водоростей еукаріотна: вона має оформлені органели з власними мембранами: ядро, мітохондрії, пластиди та ін. Зелені пігменти – хлорофіли аі з– маскуються додатковими: помаранчевими каротинами та жовто-коричневими ксантофілами; з останніх переважає фукоксантін. У клітинах бурих водоростей зустрічається від однієї до багатьох пластид. Поверх зовнішньої клітинної мембрани клітини бурих водоростей зазвичай виробляють тверду клітинну стінку.
Ядро найчастіше одне, але іноді ядер кілька. У деяких представників із збільшенням віку клітини кількість ядер збільшується. Ядро в ядрі завжди одне.
Хлоропласти вегетативних клітин бурих водоростей мають вічко (стигму) та піреноїди, яких буває один або кілька; у деяких видів піреноїди відсутні. Вічко, що складається з 30-40 ліпідних глобул, виконує функцію лінзи, що фокусує світло на джгутикове здуття, яке є фоторецептором. Зооспори та гамети бурих водоростей зазвичай не мають вічка. У піреноїдах відкладаються запасні речовини. Хлоропласти найчастіше дрібні, дископодібні, парієтальні. Їх форма може бути зірчастою, стрічковоподібною або пластинчастою. Форма хлоропластів у клітинах бурих водоростей може змінюватись у міру старіння клітини. Оболонка хлоропласту складається з чотирьох мембран, причому дві зовнішні мембрани пов'язані з ЕПС-ендоплазматичною мережею клітини.
Целюлоза, з якої складається стінка клітин бурих водоростей, її іноді називають альгулозою, за властивостями відрізняється від целюлози вищих рослин. Назовні від целюлозної стінки розташовується пектиновий шар, що сильно ослизняється, утворений в основному альгінової кислотою і її солями і сполуками з білковими речовинами. Альгінова кислотатакож виявлена лише у бурих водоростей.
Загальна характеристика.
Всі бурі водорості є фототрофами. Продукти фотосинтезу – специфічний вуглевод хризоламінарин,спирт маніті ліпіди.Ліпіди накопичуються у вигляді крапель олії. Хризоламінарин відкладається за межами хлоропласту: він розташовується на спеціальних вакуолях на цитоплазмі у піреноїду. Маніт виконує резервну, а також осморегуляторну функцію; його концентрація усередині клітини залежить від зміни солоності води.
У молодих клітинах бурих водоростей містяться дрібні та численні вакуолі, які з віком стають більшими за рахунок злиття. У цитоплазмі розташовані особливі освіти – фізиди, які містять флоротаніни.Ці речовини відомі лише у видів бурих водоростей. Функція флоротанінів, можливо, пов'язана із захистом таломів водоростей від виїдання тваринами, наприклад, молюсками. Флоротаніни бурих водоростей, ймовірно, інгібують поселення на поверхні талому епіфітних водоростей та тварин. Передбачається, що флоротаніни беруть участь у захисті від радіційного пошкодження та у виборчому поглинанні з водного середовища іонів важких металів. Безбарвні флоротаніни на повітрі окислюються з утворенням пігменту фікофеїну, який надає висушеним бурим водоростям характерного темного забарвлення.
Будова талому.Серед видів бурих водоростей відсутні одноклітинні та колоніальні форми, всі особини багатоклітинні. Форма таломів бурих водоростей найрізноманітніша. Це нитки, що стелиться або вертикально стоять, однорядні або багаторядні; масивні суцільні або черепітчасто з'єднані кірки та скоринки; тонкі слизові пластинки або щільні шкірясті пластини на стовбурі, прості або розсічені; бульбашки та мішки; трубки та шнури; по-різному розгалужені тонкі кущі або потужні кущі. Прикріплення таломів здійснюється за допомогою ризоїдів або підошви. Для утримання у вертикальному положенні у великих бентосних форм бурих водоростей утворюються повітряні (плавальні) бульбашки, заповнені газом (рис. 29).
Мал. 29. Зовнішній вигляд бурих водоростей: 1 – Фукус; 2 – Сахаріна; 3 – Аларія; 4 – Саргассум
Найбільш складно серед видів бурих водоростей влаштовані таломи представників порядків Ламінарієві та Фукусові. Вони мають ознаки тканинного диференціювання зі спеціалізацією клітин талому (рис. 30). У їх слоевищах можна розрізнити кору, що складається з інтенсивно пофарбованих клітин та серцевину, Що складається з безбарвних клітин, які часто зібрані в нитки. У Ламінарієвих у серцевині утворюються також ситоподібні трубки та трубчасті нитки. Серцевина виконує як транспортну функцію, а й механічну, оскільки у ній перебувають нитки з товстими поздовжніми стінками. Між корою і серцевиною у багатьох бурих водоростей може бути проміжний шар великих безбарвних клітин.
Зрістталому у представників бурих водоростей найчастіше інтеркалярний (вставковий) та апікальний (верхівковий), рідше базальний. Інтеркалярне зростання може бути дифузним, або є зона зростання. У великих форм вставкова (інтеркалярна) меристема, або група клітин, що активно ділиться, розташована в місці переходу «черешка» в «листову» платівку. Великі водорості також мають меристематичну зону на поверхні талому, так звану меристодерму (своєрідний аналог камбію вищих рослин).
Розмноженняу бурих водоростей вегетативне, безстатеве та статеве. Гамети формуються в багатогніздних гаметангіях, зооспори - в 
Мал. 30. Внутрішня будова Ламінарії(по: Л.Л. Великанов та ін., 1991): А- Поздовжній зріз стволика; Б– поперечний зріз стволика; У– зріз пластини із сорусами зооспорангіїв
одногніздних спорангіях.
Вегетативне розмноження у ряду видів бурих водоростей може здійснюватися ділянками талому; у видів роду Сфацелярія– вивідковими гілочками (висновковими нирками); у видів роду Фукус- групою клітин на підошві, які здатні до диференціювання у новий талом.
Безстатеве розмноження бурих водоростей зазвичай відбувається за допомогою зооспор. Деякі види розмножуються нерухомими тетраспор або моноспор. Спори безстатевого розмноження формуються в результаті мейозу і наступних мітозів одногніздних спорангіях. Статевий процес – ізогамія, гетерогамія та оогамія. Гамети утворюються в багатогнездних гаметангіях- Спеціальних групах клітин. У кожному гнізді гаметангія формується одна гамета.
Для бурих водоростей відомі статеві феромони- Розчинні речовини, які координують активність клітин при розмноженні. Феромони, яких відомо щонайменше десять, можуть або стимулювати розкриття антеридіїв, або залучати чоловічі гамети до жіночих. Вони також беруть участь у ізоляції видів. Вважають, що феромони утворюються у більшості видів бурих водоростей, що мають статевий процес.
Життєвий цикл.У бурих водоростей можна назвати два основних типи життєвих циклів. Один тип - гаплодіплобіонтний. Спорофити диплоїдні, тобто. усі клітини містять повний набір хромосом. Суперечки безстатевого розмноження формуються в одногніздних спорангіях. При утворенні суперечка відбувається редукційний поділ – мейоз. Така редукція називається споричною. Клітини (спори), що утворилися після мейозу, гаплоїдні, тобто. містять половинний набір хромосом. Гаплоїдні зооспори та тетраспори проростають у гаплоїдний гаметофіт, на якому у багатогніздних гаметангіях формуються гамети. Диплоїдна зигота, що утворилася після злиття гамет, проростає в диплоїдний спорофіт.
Інший тип – диплобіонтний; спорофіти, суперечки та гаметофіти диплоїдні; редукційний поділ відбувається при утворенні гамет. Така редукція називається гаметичною. Після злиття гаплоїдних гамет утворюється диплоїдна зигота і проростає в диплоїдний спорофіт. Цей тип поширений у видів ламінарієвих (рис. 31).

Мал. 31. Життєвий цикл Сахаріни(по: Л.Л.Великанов та інших., 1991): 1 – спорофит, 2 – сорусы зооспорангиев, 3 – зооспори , 4 – гаметофіт, 5 – антеридій, 6 – оогоній, 7 – антерозоїд, 8 –зигота (запліднена яйцеклітина)
На макроскопічному спорофіті Сахаріни, у верхній частині пластини, смугами розвиваються соруси(групи) зооспорангіїв. У зооспорангіях після редукційного поділу утворюються зооспори з двома нерівними джгутиками. Вони виростають у мікроскопічні гаметофіти: чоловічі з антеридіями та жіночі з оогоніями. Запліднена антерозоїдам яйцеклітина відразу, без періоду спокою, проростає і утворює спорофіт. На цьому життєвий цикл Сахарінизавершується.
Фукусові водорості немає спорофиов. У Фукусчоловічі та жіночі статеві органи – антеридії зі сперматозоїдами та концептакули з оогоніями та яйцеклітинами – знаходяться у поглибленнях слані – скафідіях, чоловічі та жіночі скафідії можуть перебувати на одному рецептакуліабо у різних рецептакулах. Оогонії сидять на клітці-ніжці , що виникає на стінці концептакула(Рис. 32, В, Р).Клітка оогонія зазнає трьох поділів

Мал. 32. Розмноження Фукус(по: B. Fott, 1971; W. Braune et al., 1974): A - Фрагмент талому (1 - повітряний міхур, 2 - Рецептакул); Б- Розріз через рецептакул; У– розріз жіночої скафідії; Г– оогоній; Д- Вихід яйцеклітин; Е- Розріз чоловічого скафідія; Ж– гілочка з антеридіями; 3 – вихід сперматозоїдів
ядра, перше з яких редукційне, у результаті в оогонії утворюється 8 яйцеклітин.
Антеридії зазвичай формуються на підтримують їх розгалужених нитках – парафізах(Рис. 32, Е,Ж).В результаті декількох поділів утворюється 64 сперматозоїди. Сперматозоїди Фукусмають вічко, розташоване всередині редукованого хлоропласту. У передньому кінці клітини сперматозоїда 13 мікротрубочок формують хобострум- особливу структуру, яка, як припускають, бере участь у визначенні жіночого статевого атрактанта.
Сперматозоїди залучаються до яйцеклітини Фукуснеспецифічним феромоном фукосерратеном, що виділяється яйцеклітиною. Впізнавання ж на видовому рівні пов'язане зі специфічними олігосахаридами на яйцеклітині та сперматозоїді. На поверхні яйцеклітини є різні ділянки, кожна з яких містить особливі органічні сполуки - глікопротеїни. Сперматозоїди містять глікопротеїни, організовані в ділянки на мембрані та виростах переднього джгутика, а також на поверхні сперматозоїда. При зустрічі з яйцеклітиною сперматозоїд рухається над оболонкою яйцеклітини, ніби досліджуючи її за допомогою свого переднього джгутика. Якщо глікопротеїни сперматозоїда комплементарні глікопротеїнам яйцеклітини, тільки тоді відбувається впізнавання сперматозоїду яйцеклітиною.
Систематика
Викопні знахідки, які можуть стосуватися бурих водоростей, відомі із середнього силуру (близько 425 млн. років). Але ці знахідки мають схожість і з деякими сучасними зеленими та червоними водоростями. Викопні знахідки, які можна пов'язати з сучасними бурими водоростями, відносяться до міоцену (5–25 млн. років). Ці водорості нагадують сучасну Диктіоту. Вік бурих водоростей, який визначається молекулярними методами, становить 155–200 млн. років.
Тип організації талому, наявність або відсутність піреноїду, спосіб росту, тип статевого розмноження (ізогамія, гетерогамія, оогамія) та життєвого циклу використовують для виділення порядків бурих водоростей. Останніми роками активно переглядається система бурих водоростей. У різних класифікаційних схемах у складі класу бурі водорості виділяють від 7 до 14 порядків.
Порядок Десмарестієві–Desmarestiales. Водорості кущисті, багаторазово розгалужені, з валькуватою, плоскою або сплощеною слані. Прикріплюється до субстрату добре розвиненою підошвою. На поверхні слані розвиваються волоски.
Рід Десмарестія (рис.33). Бурі або оливкові кущі з добре вираженою головною втечею, до 0.8-1 м висоти. Прикріплюється конусоподібною підошвою. Розгалуження 3–4 порядків, почергове чи супротивне. Гілки останніх порядків плоскі або, рідше, ниткоподібні. Види морські, мешкають в арктичних та помірних широтах на глибині 1-20 м, утворюють зарості або ростуть одинично.

Мал. 33. Десмарестія (Р. Desmarestia)
Порядок Ральфсієві–Ralfsiales. Представники порядку мають слоевище у вигляді кірок різної будови або складаються з базальної кірки та вертикальних пагонів.
Рід Аналіпус.Листок складається з багаторічної кіркоподібної базальної частини і однорічних, оливково-бурих, вертикальних пагонів, що відходять від неї, висотою до 40 см. Багатогніздні спорангії циліндричні за формою і дворядні за будовою. Одногніздні спорангії овальні, розвиваються серед асиміляційних гілок. Види роду широко поширені у морях помірних широт, утворюють пояси у припливно-відливній зоні.
Порядок Диктіотові–Dictyotales.Для видів порядку характерний апікальне зростання. У вегетативних клітинах є численні дископодібні хлоропласти без піреноїдів. Таллом паренхіматозний, характерна ізоморфна зміна поколінь. Статевий процес оогамний, безстатеве розмноження нерухомими спорами, сперматозоїди мають лише один джгутик. Джгутик довгий, з перистими виростами і спрямований уперед. Представники порядку зустрічаються переважно у тропічних і, рідше, в помірних морях.
Рід Диктіота (додаток, 4А). Має слоєвище у вигляді плоских оливково-бурих дихотомічно розгалужених кущиків заввишки до 11-15 см. Гілки лінійні, вузькі, не мають жилок, з роздвоєними вершинками. Тетраспорангії утворюють на поверхні слані соруси, які мають невизначену або кільцеподібну форму. Поширений у морських водах помірних та теплих широт.
Порядок Ламінарієві–Laminariales. Ламінарієві мають слоевище, що складається з пластини, стволика та органів прикріплення – підошви або різозідів. Пластина рівна або зморшкувата, цільна або розсічена, з отворами, складками, булями або без них, завдовжки від кількох десятків сантиметрів до 20-40 м. Стволик розгалужений або частіше нерозгалужений. Спорофити однорічні або багаторічні, вік деяких видів налічує 11-18 років. У корі багатьох ламінарієвих розташовані слизові оболонки.
Найбільш популярними представниками бурих водоростей є ламінарії (види пологів Ламінарія або Сахаріна), або «морська капуста», які поширені переважно у Тихому океані. Вони зустрічаються від поверхні (урізу) води до глибини 200 м. Зростають ламінарії переважно на глибині 2–20 (30) м, у місцях із постійним рухом води, прикріплюючись до каменів та скель. Ламінарієві утворюють водоростеві пояси вздовж узбережжя морів Далекого Сходу. Найбільш потужні зарості зустрічаються біля Курильських островів і в о. Хоккайдо, де сконцентровано близько половини світових ресурсів ламінарієвих. Щорічний світовий збір ламінарієвих водоростей сягає 2 млн. тонн сирої маси, причому понад 1 млн. тонн дає виробництво її марикультури в Китаї.
Рід Сахаріна (Ламінарія).Листок довжиною від 0,5 до 5-10 м, має товсту блискучу пластину, пружний стволик і густо переплетені різоїди, якими прикріплюється до різних твердих субстратів. Краї пластини гладкі або хвилясті, на поверхні пластини часто є опуклості та складки. Скупчення спорангіїв – соруси – утворюються по обидва боки пластини, після виходу суперечка верхня частина пластини руйнується. Живе від двох до кількох років. Сахаріни є цінною промисловою сировиною та об'єктом марікультури. Сахаріна японськаутворює зарості у північній половині Японського моря, а також біля південного та південно-східного берега Сахаліну та біля Південних Курильських островів. Сахаріна японськає найбільш цінним промисловим видом, вона відрізняється чудовими смаковими якостями та використовується для приготування різноманітних страв, консервів, кондитерських виробів.
Порядок Сфацелярієві–Sphacelariales.
Бурі водорості, що відносяться до цього порядку, мають таллом, що гілкується, з апікальним зростанням. Гілки з волосками. Вегетативні клітини мають численні хлоропласти без піреноїдів. Життєвий цикл у більшості з ізоморфною зміною поколінь, статевий процес – ізогамія або гетерогамія. Водорості утворюють невеликі бурі пучки до 3 см у висоту та розгалужені кущики до 20 см у висоту на камінні або інших водоростях; є ендофітні представники.
Рід Сфацелярія. Має тонконітчасте слоевище. Нитки, що гілкуються, утворюють невеликі, до 0,5-3 см висоти, пучочки бурого кольору. Вегетативне розмноження здійснюється спеціалізованими, часто трипроменевими вивідковими гілочками. пропагулами. Статевий процес ізогамний. Жіночі гамети виділяють феромони ектокарпен та мультифіден. Види роду поширені у морях як полярних, і тропічних широт.
Порядок Сцитосифонові–Scytosiphonales. Представники з кишкоподібною, міхуровою, пластинчастою або мішковидною, часто порожнистою слані до 25 см довжини. Кора однорядна або дворядна, внутрішня частина (стінка) слані утворена кількома шарами великих клітин. На поверхні слані розвиваються багатогніздні спорангії, які зібрані в соруси, що підтримують їх нитки. парафізита пучки волосків.
Рід Сцитосифонє тонкостінними бурими трубками 4–25 см довжини (додаток, 4Б). Листок прикріплюється невеликою підошвою і має рідкісні поперечні перетяжки. Зустрічається у прибережній зоні морів у різних широтах, часто утворює пояси у припливно-відливній зоні.
Порядок Фукусові–Fucales. Мають багаторічну слоевище у вигляді великих кущів, що неправильно або дихотомічно гілкуються, до 1-3 м довжини. Прикріплюються до субстрату конусоподібною підошвою. Слані мають пагони, філоїди (листкоподібні утворення), рецептакули (містилища гамет) і плавальні (повітряні) бульбашки. Зустрічаються в припливно-відливній зоні всіх помірних та арктичних морів, де утворюють потужні чагарники – пояси.
Рід Саргассум(Рис. 34, У). Рясно розгалужені оливково-бурі або коричневі кущі заввишки 05-15 м. Прикріплюються конічною підошвою. Філоїди розвиваються на бічних пагонах, що відходять від циліндричного стовбура, в нижній та середній частинах слані. Філоїди шкірясті, з центральним ребром, 0,2-2 см довжини. Плавальні пухирі овальні або округлі, з виростами або без них. Саргасси широко поширені в помірних та субтропічних широтах від нижньої літоралі до глибини 10 м, де утворюють щільні або розріджені чагарники. У південній частині Тихого океану – так званому Саргасовому морі – утворюють великі плаваючі скупчення, у яких розмножуються лише вегетативним шляхом.
Рід Фукус (рис.32).Словище шкірясте, кущисте, розгалужується дихотомічно, висотою 25-50 см. Біля узбережжя далекосхідних морів росте кілька років, мешкає на літоралі. Часто утворює великі зарості на мілководді. Для видів цього роду характерно дихотомічно

Мал. 34. Зовнішній вигляд фукусових водоростей: А – Аскофілум; Б – Цистозіра; У – Саргассум; 1 – рецептакул, 2 – плавальний міхур
розгалужена слоевище з плоскими гілками, що мають поздовжнє ребро, яке прикріплюється до каменів конічної підошвою. Фукуси використовують як добрива, як корм для худоби, для виробництва кормового борошна, альгінатів, застосовують у медицині для лікування зоба, корекції ваги і т.д.
Порядок Ектокарпові–Ectocarpales. Водорості, які сюди належать, мікроскопічні, вільноживучі, епіфіти, епізоїти чи ендофіти. Органи розмноження представлені одногніздними (спорангії) та багатогніздними (гаметангії) спорангіями; вони довгоциліндричні, короткоциліндричні або стручкоподібні.
Рід Ектокарпус. Лист у вигляді м'яких бурих кущиків до 2 см висоти. Кущики утворені однорядними, по черзі, дихотомічно або неправильно розгалуженими нитками. Багатогніздні спорангії стручкоподібні, утворюються збоку бічних гілок на 1-3-клітинній ніжці. Види поширені в морях різних широт, у чистих та забруднених водах, на літоралі та у верхній частині субліторалі. Епіфіти зустрічаються і в обростанні антропогенних субстратів.
Екологія
Переважна більшість представників бурих водоростей живе у морських водоймах. У прісних водах трапляються лише 8 видів. Трапляються як однорічні, так і багаторічні види, вік яких може досягати 15-18 років. Ламінарієві водорості Нереоцистісі Макроцистис, Які мешкають вздовж тихоокеанського узбережжя Північної Америки, мають максимальні для всіх водоростей розміри - до 30-40 м завдовжки. Вони формують гігантські підводні ліси морях Далекого Сходу. Прикріплюються бурі водорості до найрізноманітніших субстратів – скель, каменів, гравію, раковин та панцирів морських тварин, інших водоростей. Деякі дрібні форми бурих водоростей живуть усередині тканин інших водоростей як ендофіти. Більшість видів бурих живуть у прикріпленому стані. Відірвані від ґрунту слані зносяться течією у спокійні місця з мулистим дном і продовжують там існувати. Види з повітряними бульбашками на слоевище, відірвавшись від ґрунту, спливають до поверхні, утворюючи великі плаваючі скупчення, особливо в районах зі стабільною круговою течією, як у Саргасовому морі. Вони існують у таких скупченнях досить довгий час і розмножуються лише вегетативним способом.
Бурі водорості широко поширені у всіх морях, але найбільшого розвитку вони досягають у морях помірних та приполярних широт. Їхні великі слані ростуть, прикріпившись до скель і каменів, а в спокійних місцях біля берега і на великій глибині можуть утримуватися навіть на стулках раковин молюсків та гравії. Зустріти їх можна на різній глибині – від літоральної зони, де під час відливу вони годинами знаходяться поза водою до 40–200 м. У районі Гавайських островів на глибині 180 м виявлено види роду Саргассум, а Ламінаріяв Адріатичному морі було знайдено на глибині 200 м. Найбільш рясні зарості бурих водоростей спостерігаються на глибині 6–15 м, де найкращі умови освітлення та постійний рух води, що приносить до їх слоїв біогенні речовини та обмежує поселення рослиноїдних тварин. фітофагів.
Золотисті водорості –Chrysophyceae
Одноклітинні, колоніальні або багатоклітинні організми, які зазвичай мають джгутикову стадію в життєвому циклі. Клітини одноядерні, з одним або декількома хлоропластами. Стигма зазвичай є (рис. 36). Жгутиків 1-2, нерівних. Скорочувальна вакуоль розташована в передній частині клітини. Зовнішні покриви клітин цих організмів дуже різноманітні. У найбільш примітивних видів клітина одягнена ніжним перипластом, що дозволяє утворювати випинання різної форми (ризоподії, псевдоподії), в інших плазмалемму покриває тверда целюлозна оболонка, що іноді сильно ослизняється. Будиночки у золотистих водоростей бувають різної форми: вазоподібні, кулясті, яйцеподібні, циліндричні; з одним або декількома отворами. Клітина прикріплюється до основи будиночка за допомогою гнучкої ніжки або може бути вільною. У деяких хризофіт на поверхні клітини відкладаються вапняні утворення - коколіти - різних форм і розмірів, а кремнієві джгутиконосці мають внутрішній кремнієвий скелет. Різноманітність будови цих скелетів дозволяє використовувати залишки тих чи інших форм викопних силікофлагелят для визначення віку геологічних порід, що їх містять. У більш високорозвинених представників відділу золотистих водоростей клітини одягнені панциром, що складається з кремнієвих лусочок (іноді несучі шипи), або укладені в будиночки, через отвори яких назовні виходять джгутики або псевдоподії.
Будиночки у видів золотистих водоростей вазоподібні, кулясті, яйцеподібні, циліндричні, з одним або декількома отворами. Клітина прикріплюється до основи будиночка за допомогою гнучкої ніжки або може бути вільною.
Пігменти – хлорофіли аі з, фукоксантін. Для хризофіт характерний найвищий серед гетероконтів вміст фукоксантину (55-93% від усіх каротиноїдів).
Більшість видів - прісноводні фототрофи, але зустрічаються гетеротрофи та фаготрофи.
Запасні речовини – хризоламінарин та олії.
Серед одноклітинних форм золотистих водоростей переважають прісноводні планктонні види, серед багатоклітинних – донні чи эпифитные. Зустрічаються в основному в прохолодних морських та прісних водоймах. Відомо близько 360 видів.

Мал. 36. Золотисті водорості (по: С. Hoek van den et al., 1995): А– будова клітини; Б- Корінецька система; У- Тричастинна мастигонема; Г- Зріз частини хлоропласту; 1 – базальне тіло; 2 – базальне здуття; 3 – короткий джгутик; 4 – стигма; 5 – ризопласт; 6 – ядро; 7 – ядерна мембрана; 8 - ядерце; 9 – мітохондрія; 10 – вакуолі; 11 - слиз; 12 – плазмалема; 13 - везикули; 14 – слизове тіло; 15 – ліпіди; 16 – хлоропласт; 17 - апарат Гольджі; 18 – скорочувальна вакуоля; 19 - довгий джгутик: 20 - 1-й корінець; 21 - 2-й корінець; 22 - 3-й корінець; 23 - 4-й корінець; 24 - хлоропластний нуклеоїд; 25 - ХЕС; 26 – оболонка хлоропласту; 27 - оперізує ламелла; 28 - ламелла; 29 – довгий латеральний філамент; 30 – короткий латеральний філамент; 31 – термінальний філамент; 32 - трубчаста частина мастигонеми
Структура таломуу золотистих водоростей найчастіше монадна, але може бути дуже різноманітною: кокоїдною, пальмелоїдною, плазмоїдальною, амебоїдною, нитчастою, паренхіматозною (рис. 37).

Мал. 37. Золотисті водорості (по: Л.Л. Великанов та ін, 1981): А – Охромонас: 1 – зовнішній вигляд; 2 – циста; Б – Хромуліна: 1 – зовнішній вигляд; 2 – циста; 3 – схема утворення плівки з цист; У – Дінобріон: 1 – загальний вигляд колонії; 2 – циста; Г – Хризамеба; Д – Гіббердія; Е – Гідрурус: 1 – загальний вигляд колонії, 2 – кінчик гілки, 3 – зооспора
Розмноження. У золотистих водоростей зустрічаються три способи розмноження: вегетативне, безстатеве і статеве.
Вегетативне розмноженнявідбувається шляхом поздовжнього поділу клітини або розпаду колонії на частини за рахунок поділу клітин в одному, двох або трьох напрямках.
Статевий процес(клітини золотистих водоростей диплоїдні) - ізогамія або кон'югація. В результаті статевого процесу, а також у несприятливих умовах утворюються цисти з товстою оболонкою, що містить кремній.
Безстатеве розмноженняздійснюється одно- або двожгутиковими зооспорами, які розвиваються у звичайних вегетативних клітинах або зооспорангіях. Статевий процес: голо-, ізо- та автогамія. Гологамія – вид статевого процесу, при якому дві подібні з вегетативними клітинами зливаються на передніх кінцях у двоядерну зиготу, яка перетворюється потім на окремі цисти.
Систематика
Із Золотистих водоростей наприкінці минулого століття на основі результатів молекулярно-генетичних досліджень виділено клас Синурові. Також виділено клас Феотамнієві – із Жовтозелених та Золотистих водоростей. Тому обсяг класу Chrysophyceae скоротився і в даний час налічує близько 360 видів, що належать до трьох порядків: Chromulinales, Hibberdiales і Hydrurales. Форма, будова та кількість джгутиків, а також пігментний склад та структура тіла є важливими систематичними ознаками.
Порядок Хромулінові – Chromulinales.
Рід ХризамебаДо цього порядку відносять організми з монадним, пальмелоїдним та амебоїдним типами диференціації талому. Монадні клітини з одним джгутиком, видимим у світловий мікроскоп. А(Рис. 37,
Рід ХромулінаДо цього порядку відносять організми з монадним, пальмелоїдним та амебоїдним типами диференціації талому. Монадні клітини з одним джгутиком, видимим у світловий мікроскоп. У) – прісноводні амебоїдні водорості, вони мають у життєвому циклі джгутикову стадію. Зигота перетворюється на стоматоцисту.
Рід Дінобріон) - одноклітинні вільноплаваючі організми, що мешкають переважно в прісних водах. Клітини голі; є одна чи дві золотисті пластиди. (Рис. 37, Г)
– одноклітинні та колоніальні, вільноплаваючі або прикріплені представники. Монади знаходяться у вазоподібних будиночках, утворених целюлозними мікрофібрилами. Крім целюлози до складу будиночка входить і багато амінокислот. Кущикоподібна форма колоній пов'язана зі способом вегетативного поділу клітин, коли одна з дочірніх клітин, залишаючи батьківський будиночок, прикріплюється до його отвору. Обертаючись навколо своєї осі, вона формує власний будиночок. Батьківський будиночок можуть залишити і обидві дочірні клітки. Клітини містять 1-2 хлоропласти, на передньому кінці клітини видно вічко і 2 скорочувальні вакуолі. Хризоламінарин розташований у вакуолі на задньому кінці клітини. Розмноження вегетативне та статеве.–Порядок Гіббердієві Hibberdiales
. Для представників цього порядку характерний унікальний пігментний склад. Крім фукоксантину, вони мають додатковий світлозбираючий каротиноїдний пігмент - антераксантин. Включає пологиГіббердія, Хромофітон, Стилоцерас, Хризопіксис, Платитека
Рід Гіббердіята інші.
має дві стадії в життєвому циклі: колоніальну пальмелоїдну нерухому і одноклітинну рухливу монода. При світловій мікроскопії у монадної стадії видно лише один джгутик. Характерна наявність крім фукоксантину ще одного додаткового світлозбираючого каротиноїдного пігменту – антераксантину.–Порядок Гідрурусові. Представники даного порядку мають таллом з пальмелоїдним та псевдопаренхіматозним типами будови. Характерна наявність унікальних зооспор тетраедричної форми. Ока немає. Мітоз напівзакритий.
. Для представників цього порядку характерний унікальний пігментний склад. Крім фукоксантину, вони мають додатковий світлозбираючий каротиноїдний пігмент - антераксантин. Гідрурус, Целлонієлла, Федерматіум, Хрізонебулата інші.
Рід Гідрурус(Рис. 37, Е) є найбільш диференційованим серед пальмелоїдних водоростей. Його талом є великими, довжиною до 30 см слизові колонії, що мають вигляд коричневих розгалужених шнурів, що часто видають дуже неприємний запах. У колоніях можна розрізнити головний стовбур та бічні розгалуження. У клітинах, занурених у загальний слиз, міститься один чашоподібний хроматофор та кілька скорочувальних вакуолей. Клітини рихло розташовані по периферії колонії і щільніше в середині. Таллом може зростати лише за рахунок поділу апікальних клітин. При безстатевому розмноженні у клітинах бічних гілок колонії формуються зооспори. Утворює кулясті цисти. Зустрічається у гірських струмках та річках з холодною водою, де прикріплюється до твердих субстратів.
Клас Синурові водорості –Sinurophyceae
syn- разом і ura– хвіст. Ця група організмів у 1987 р. виділена із золотистих водоростей. Поєднує монадні одиночні та колоніальні організми, іноді з чергуванням у життєвому циклі монадної та пальмелоїдної стадії. Поверхня пектинових оболонок покрита панциром із кремнієвих лусочок. Мітохондрії з трубчастими христами зазвичай розташовані в цитоплазмі навколо хлоропласту. Ядро одне, хлоропластів, оточених чотирма мембранами, зазвичай два. Ламели тритілакоїдні, оперізуюча ламела є. Основні пігменти – хлорофіли аі з, β -каротин та фукоксантин. Стигми немає. Клітини зазвичай з 2 нерівними джгутиками. Довгий перистий джгутик спрямований уперед. Короткий гладкий джгутик, який іноді сильно редукований, спрямований назад.
Клітини розмножуються переважно поздовжнім розподілом. Колонії, розпадаючись, дають початок молодим колоніям. У деяких видів описаний статевий процес у формі ізогамії. При цьому, на відміну від золотистих водоростей, злиття ізогамет у синурових водоростей відбувається не на передніх, а на їх задніх кінцях. Ендогенним шляхом утворюються окремі цисти з часом, подібні з такими у хризофіт.
Переважно планктонні форми, пальмелоїдні стадії входять до складу бентосу. Більшість видів синурових водоростей – фототрофні прісноводні організми.
Систематика.
Клас синурових водоростей включає 7 порядків: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales і Thallochrysidales.
Порядок Синурові – Synurales. Монадні форми, пектинові оболонки клітин зазвичай покриті панциром з кремнеземних лусочок, зцементованих органічною речовиною в цілісний футляр. Хлоропластів один чи два.
Рід Синура(Рис. 38) - прісноводні монадні форми з двома нерівними джгутиками та двома пристінковими хлоропластами. Ядро грушоподібне, розташоване у передній частині клітини. Позаду ядра є одна велика вакуоль, а задній частині клітини – кілька дрібних скорочувальних вакуолей. Клітини покриті лусочками, як черепицею Розмноження найчастіше вегетативне, для деяких відомий статевий процес. Утворює колонії (рис. 39, А). Часто у всіх клітинах колонії виникають цисти. При масовому розвитку Синураможе надавати воді неприємного запаху.

Мал. 38. Зовнішній вигляд клітин Синури
Порядок Ризохлоридові – Rhizochloridales. Переважно прісноводні організми з ризоподіальним типом диференціації талому, що живуть головним чином у прісних водоймах.
Рід Малломонас(Рис. 39, Б-Г) – одноклітинна монада з одним добре помітним джгутиком та одним роздвоєним хлоропластом.

Мал. 39. Зовнішній вигляд синурових водоростей (по: Г.А. Белякова та ін., 2006): А – колонія, що ділиться Синури; Б, В – освіта статоспори та Г – зовнішній вигляд Малломонаса
Клітина покрита лусочками черепітчасто і по спіралі, деякі з них несуть голкоподібні шипи (рис. 39, Г). Характерна наявність статоспор (рис. 39, Б, В). Для низки представників описаний статевий процес – гологамія.
Рід Міксохлорісмешкає в клітинах листя сфагнових мохів, має вигляд великого багатоядерного плазмодія. Восени він утворює цисти, які проростають навесні. З них виходять зооспори або амеби, що проникають у порожні клітини листя і зливаються там у плазмодії.
Порядок хлоромебових - Chloramoebales.Включає монадних представників. Зустрічаються у солоних та прісних водах.
Рід Гетерохлоріс – його клітини можуть змінювати форму, утворюючи псевдоподію. Це явище притаманне багатьом представникам порядку, як і тенденція до тваринного способу харчування. У клітці є кілька хлоропластів, краплі олії та хризоламінарин. У передній частині клітини є скорочувальні вакуолі, у центрі – одне ядро. Розмножується вегетативно поздовжнім поділом клітини.
Порядок Охромонадові - Ochromonadales. Поєднує голі форми з двома нерівними джгутиками. Прісноводні та морські форми.
Рід Охромонасвключає одноклітинні монади з двома нерівними джгутиками. Клітини одягнені лише плазмалемою. На передньому кінці клітини є скорочувальна вакуоля та вічко, на задньому кінці – вакуоля з хризоламінарином. Розмножується вегетативно, борозна розподілу починається біля переднього кінця клітини між двома парами джгутикових основ. Види роду поширені в оліготрофних прісних водах, але є і морські представники.
Порядок Гетероглеєві - Heterogloeales. Включає водорості з пальмелоїдним типом диференціації талому. Представники порядку зустрічаються частіше у прісних, ніж у солоних водах.
Рід Гельмінтоглеямешкає в солонуватих водах. Ця колонія, що сидить на розширеній підставі, складається з слизових тяжів, що гілкуються. У цьому слизу безладно розташовуються протопласти, кожен із яких оточений власної слизової обгорткою.
Клас Феотамнієві водорості – Phaeothamniophyceae
Назва класу походить від типового роду Phaeothamnion(Від грец. phaeos- Смаглявий кущик). Представників цієї групи організмів виділили в окремий клас із жовтозелених та золотистих водоростей у 1998 р. на підставі аналізу послідовності рибосомальних генів та особливостей ультраструктури клітини. Для феотамнієвих водоростей характерна унікальна комбінація пігментів: фукоксантину з гетероксантином, відсутність віолаксантину.
Особливості будови клітини: відсутність вакуолей із хризоламінарином та відсутність ендогенних цист із кремнеземними стінками.
Клас включає одноклітинні, колоніальні та багатоклітинні організми з кокоідним, пальмелоїдним та нитчастим типом структури талому. Жгутикові стадії з двома нерівними джгутиками. Джгутики латеральні або субапікальні. Стигма зустрічається у зооспорі. Пігменти – хлорофіли аі з, β -каротин, фукоксантин, діадіноксантін, діатоксантин та гетероксантин. Основним запасним продуктом є парамілон ( β -Глюкан). Клітинна стінка є, під час клітинного поділу дочірня стінка формується усередині батьківської стінки.
У феотамнієвих з основних способів розмноження зустрічається лише вегетативне та безстатеве. Безстатеве розмноження здійснюється автоспорами або зооспорами. Статеве розмноження невідоме.
Види Феотамнієвих водоростей мешкають виключно у прісних водах. Систематика.
В даний час підтверджена монофілетичність класу феотамнієвих водоростей, і серед охрофіту він ближчий бурим і жовтозеленим водоростям, ніж золотистим. В даний час відомо близько 30 видів цих водоростей, що належать до одного порядку Феотамнієві – Phaeothamniales.
Рід Феотамніонявляє собою прикріплені нитки, що гілкуються, до 1 см висоти (рис. 40). Клітини циліндричні, неправильні, розширені догори,

Мал. 40. Зовнішній вигляд Феотамніону.
довжині вдвічі-втричі перевищують ширину. При розподілі клітин матеріал батьківської стінки витрачається на утворення шаруватого чохла, що оточує клітини нитки. По периферії клітини під плазмалемою є везикули. Вони нагадують фізиди – утворення, що зустрічаються у клітинах бурих водоростей. Клітини містять від однієї до кількох дископодібних пластид оливково-бурого кольору. Зооспори утворюються по 1-2, рідше по 4-8 в одній клітині. У зооспор джгутики прикріплюються збоку. Положення джгутикових корінців у Феотамніонунагадує таке у жовтозелених та бурих водоростей. У нього також відсутня ризопласт. У життєвому циклі можуть утворюватися цисти, але у стінках відсутня кремній. Цисти проростають зооспорами. Поселяється як епіфіт на ниткових водоростях. Мешкає у стоячих та повільно поточних прісних водоймах.
Клас Рафідові водорості –Raphidophyceae
Назва класу походить від грец. rhaphid - Голка. Поєднує одноклітинні двожгутикові організми, позбавлені клітинної стінки. В основному прісноводні організми із сплощеними клітинами (рис. 41).
Клітини мають джгутики нерівної довжини, спрямований вперед джгутик - перистий і довше, спрямований назад - гладкий і коротший. Перехідна спіраль відсутня. Хлоропласти дрібні, численні чи, рідше, по 1-2 у клітині, оточені чотирма мембранами, їх дві мембрани ХЕС. Тилакоїди зібрані в стоси по три. Ядро велике, оточене кільцем диктіосом. Мітохондрії з трубчастими христами. Є скорочувальні вакуолі. Цитоплазма часто буває вакуолізованою. Клітини голі, оточені лише плазмалемою. Стигми (вічка) зазвичай немає.
Фотосинтетичні пігменти – хлорофілі аі з, β -каротин, вошеріаксантін, діноксантін, діадіноксантін, гетероксантин.
Запасні продукти асиміляції – жири та олії, рідше крохмаль та хризоламінарин.
Мешкають також у сфагнових болотах та інших місцеперебуваннях з кислою та нейтральною реакцією води. Деякі види зустрічаються в солонуватих та морських водах.

Мал. 41. Зовнішній вигляд рафідової водорості.
Систематика.
Дуже довго цю групу водоростей через зелене забарвлення, наявність глотки та ряд інших особливостей будови розглядали як клас Хлоромонади серед Євгленових водоростей. Але дані цитології, біохімії, фізіології та молекулярної біології показали їхню безперечну приналежність до відділу Гетероконтні (Охрофітові) водорості та монофілетичність групи Рафідових водоростей.
Відомо близько 25 видів з єдиного порядку Хаттонелові– Chattonellales.
Рід Гетеросигма(Рис. 42, А) включає морські флагеллати. Клітини овальної форми зі джгутиками, зрушеними на бік, пластиди розташовані по периферії клітини. Трихоцист немає. Плавають, здійснюючи обертові рухи навколо поздовжньої осі тіла. Можуть викликати токсичне "цвітіння води" у прибережних водах.
Мал. 42. Рафідові водорості (по: R. E. Lee, 1999): А – Гетеросигма; Б – Гоніостомум; 1 – хлоропласт; 2 – мукоциста; 3 – скорочувальна вакуоля; 4 – трихоциста; 5 – ядро
Рід Гоніостомум(Рис. 42, Б) - рухливі дорсовентрально стислі монади. Спинна сторона вигнута, черевна – сплощена. Борозна на передньому кінці веде в трикутну горлянку, з якої виходять два довгі джгутики; їх довжина можна порівняти з довжиною клітини. Пластиди розташовані на периферії клітини. Під плазмалемою знаходяться трихоцисти. Мешкає в прісних водах із кислотним значенням рН.
Клас Евстигмові водорості –Eustigmatophyceae
Назва класу походить від грец. eu- "добре та stigma-«Мітка», «пляма». Клас поєднує голі одноклітинні, рідше колоніальні, організми переважно кокоідної структури.
Ядро одне чи кілька. Хлоропласт зазвичай один, оточений 4 мембранами. Джгутик зазвичай один передній перистий і друге базальне тіло або, рідше, два нерівні джгутики. Стигма (вічко) є, розташована за межами хлоропласту, її гранули без мембран.

Мал. 43. Зовнішній вигляд евстигмової водорості
Хлоропласт зазвичай єдиний, великий, багатолопатевий, чашоподібний або парієтальний, жовто-зеленого кольору. Тилакоїди зібрані в ламелли по три, оперізувальна ламелла відсутня.
Основні пігменти: хлорофіла, β-каротин, віолаксантин і вошеріаксантин. Піреноїд зазвичай є тільки у вегетативних форм.
Запасні речовини – олії та речовина невстановленої природи, яка відкладається у вигляді твердого матеріалу поза пластидами. Крохмаль не утворюється.
Мешкають у прісних водах, рідше зустрічаються в морях та у ґрунті.
Систематика.
Раніше цю групу організмів відносили до Жовтозелених водоростей. Нині Євстигмові водорості розглядають у ранзі класу у відділі Гетероконтофіти. Відомо близько 35 видів, переважно прісноводних, що належать до єдиного порядку Євстигмові – Eustigmatales.
Рід Євстигматос(Рис. 43) - одноклітинні дрібні кулясті водорості. Клітинна стінка гладка, цілісна, без орнаменту. Жовто-зелений хлоропласт один, лопатевий, парієтальний, зі стебловими багатогранними піреноїдами. Велика центральна вакуоля з червоним вмістом. Розмножується 2 або 4 автоспорами або бутілеподібними зооспорами з одним вхідним джгутиком, великою стигмою, розташованою за межами хлоропласту на передньому кінці, і єдиним заднім хлоропластом без піреноїду. Звичайний компонент ґрунтових флор. Відомий з Нової Зеландії, Австрії, Ісландії, та з басейнів Арізони як "гірчична водорість".
Клас Жовтозелені водорості –Xantophyceae
До Жовтозелених відносяться водорості, хлоропласти яких забарвлені у світло-або темно-жовтий колір, дуже рідко зелений і лише іноді блакитний. Забарвлення таломів визначається наявністю в хлоропластах клітин наступних пігментів – хлорофілів аі з, β -каротину та ксантофілів. Переважна більшість останніх і зумовлює своєрідність забарвлення жовтозелених водоростей. Крім того, в клітинах як основний продукт асиміляції накопичуються парамілон, краплі олії і лише у деяких видів, крім того, глибки лейкозину і волютину. Крохмаль у жовтозелених водоростей не утворюється. Відмінною особливістю жовтозелених є наявність у вегетативних клітин монадної структури та у зооспор двох нерівних джгутиків. До складу клітинної стінки входять целюлоза, глюкоза та уронові кислоти. Клітинна стінка часто складається із двох частин.
Розмноження вегетативне, безстатеве та статеве.
Широко поширені у прісних водах. Рідко зустрічаються в морських, солонуватих водах та у ґрунті.
Раніше клас Жовтозелених водоростей називався Трібофіцієві водорості за типовим родом Трібонема (від грец. tribon – майстерний, хитрий і nema – нитку). Відомо близько 450 видів.
Жовтозелені характеризуються значною морфологічною різноманітністю. Серед численних представників цього відділу виявляються майже всі основні типи структури тіла: амебоїдна, монадна, пальмелоїдна, кокоїдна, нитчаста, різнонітчаста, пластинчаста та сифональна (рис. 44). – 46). Словник одноклітинний,

Мал. 44. Зовнішній вигляд жовто-зелених водоростей: 1, 2 – Хараціопсис, 3 – Центритрактус, 4 – Офіоцитіум
колоніальне, багатоклітинне та неклітинне. Клітинна оболонка щільна, пектинова і целюлозна, що складається з частин, що щільно налягають один на одного, або з двох стулок. В оболонці відкладається кремнезем чи вапно. Переважно нерухомі форми. Серед одноклітинних видів є рухомі форми, позбавлені щільної оболонки та забезпечені джгутиками, лобоподіями та різоподіями.

Мал. 45. Зовнішній вигляд ксантококкових жовто-зелених водоростей: 1–3 – Ботрідіопсис, 4 – Тетраедрієлла, 5 – Псевдостаураструм, 6 – Гоніохлоріс, 7, 8 – Бумілеріопсис
Більшість жовтозелених – нерухомі організми. У рухомих особин рух може здійснюватися за допомогою джгутиків чи різоподій. Клітини різної форми: кулясті, веретеноподібні, еліпсоїдні, циліндричні, тетраедричні, серпоподібні, грушоподібні, яйцеподібні. Талломи розмірами від 0.5 – 1.5 мкм ( Хлорідела) до кількох міліметрів у діаметрі ( Ботрідіопсис) (рис. 45, 1 – 3) і до десятків сантиметрів завдовжки ( Вошерія) (рис. 46, 3).

Мал. 46. Зовнішній вигляд жовто-зелених водоростей: 1 – Трибонема, 2 – Гетеропедія, 3 – Вошерія, частина нитки з оогонієм та антеридієм
Більшість видів жовтозелених є фототрофами, але трапляється і голозойне харчування шляхом заковтування бактерій та дрібних водоростей. Жовтозелені водорості широко поширені у прісних водах. Вони також звичайні в ґрунті, рідше зустрічаються в морських та солонуватоводних водоймах. Клас включає аеробіонтні, планктонні, донні та перифітонні форми. Епіфіти, епізоїти, а також внутрішньоклітинні симбіонти у найпростіших клітинах.
Незалежно від зовнішньої структури, внутрішня будова клітини жовто-зелених водоростей однотипна. У протопласті спостерігається зазвичай кілька жовто-зелених хлоропластів, що мають дископодібну, коритоподібну, пластинчасту, рідше стрічкоподібну, зірчасту або чашоподібну форму з цільними або лопатевими краями. Забарвлення пов'язане з відсутністю фукоксантину, що відповідає за золотистий та коричневий колір в інших охрофітових. З інших пігментів вони мають β -каротин, вошеріаксантин, діатоксантин, діадіноксантін, гетероксантин. У рухомих форм на передньому кінці хлоропласту зазвичай розташоване червоне вічко, або стигма. У небагатьох видів є піреноїди напівзавантаженого типу. Ядро в клітині зазвичай одне, невеликих розмірів, але є види і з багатоядерними клітинами. У деяких видів у передній частині клітини є одна або дві скорочувальні (пульсуючі) вакуолі.
У монадних представників і рухомих стадій (зооспор і гамет) є два нерівних джгутика. Винятком є синзооспори Вошерії, у яких по поверхні розташовані численні пари трохи розрізняються по довжині гладких джгутиків. Короткий джгутик закінчується акронемою. Джгутики прикріплюються на клітині субапікально. У сперматозоїда Вошеріїприкріплення латеральне.
У видів з амебоїдною, монадною та пальмелоїдною організацією клітинна стінка відсутня, вони покриті лише цитоплазматичною мембраною і можуть легко змінювати форму. Іноді «голі» клітини знаходяться усередині будиночків, стінки яких можуть бути забарвлені у бурий колір солями марганцю та заліза. У більшості форм є клітинна стінка, що складається з двох частин. У складі клітинної стінки переважає целюлоза, а також містяться полісахариди, що складаються в основному з глюкози та уронових кислот. У молодих клітин оболонка тонка, з віком потовщується. У ній можуть відкладатися солі заліза, з'єднання якого забарвлюють її у різні відтінки коричневого та червоного тонів. Найчастіше в клітинній стінці є кремнезем, надаючи їй твердість і блиск. Вона може інкрустуватися також вапном і бути по-різному скульптурована (шипики, осередки, бородавки, щетинки, зубчики і т.д.) У прикріплених форм може утворюватися виріст оболонки – ніжка з прикріпною підошвою.
У нитчастих форм жовтозелених водоростей із двостулковими оболонками при розпаді ниток клітинні оболонки розвалюються на Н-подібні фрагменти. Ці фрагменти є щільно з'єднаними половинками оболонок двох сусідніх клітин (рис. 47). При зростанні ниток Н-подібний фрагмент клітинної стінки двох сусідніх дочірніх клітин вбудовується між двома половинками материнської оболонки клітини. У результаті кожна з дочірніх клітин наполовину покрита старою оболонкою материнської клітини та наполовину. – новоствореною оболонкою.

Мал. 47. Схема утворення поперечної перегородки між двома дочірніми клітинами у нитчастих жовто-зелених водоростей (по: А.А. Масюк, 1993): А- Фрагмент нитки; Б- Закладка пояскового кільця та утворення поперечної перегородки між двома клітинами; У– шаруватість двостулкових клітинних оболонок; Г- Розпад оболонки на Н-подібні ділянки
Скорочувальні вакуолі є у рухливих представників. Зазвичай їх 1-2 на клітину. Апарат Гольджі своєрідної будови. Диктіосоми дрібні, містять 3-7 цистерн. Ядро одне, рідше їх багато; у ценотичних видів клітини завжди багатоядерні.
Розмноження. Для більшості видів жовтозелених водоростей характерно вегетативне та безстатеве розмноження.
Вегетативне розмноженняздійснюється різними способами: розподілом клітин навпіл, розпадом колоній та багатоклітинних слоевищ на частини. У Вошеріїутворюються спеціальні виводкові нирки.
При безстатевому розмноженніможуть формуватися різноманітні суперечки: амебоїди, зооспори, синзооспори, автоспори, гемізооспори, геміавтоспори, апланоспори. Зооспори «голі» і зазвичай грушоподібної форми.
Статевий процес– ізогамія, гетерогамія та оогамія – описаний у небагатьох представників. У Трибонемигамети подібні за розмірами, але різняться поведінкою – це ізогамія. У Вошеріїспостерігається оогамія: на нитках утворюються вмістилища жіночих гамет – оогонії та чоловічих – антеридії.
У несприятливих умовах спостерігається утворення цистів. Цисти (статоспори) ендогенні, одноядерні, рідше багатоядерні. Їхня стінка часто містить кремнезем і складається з двох нерівних або, рідше, рівних частин.
Систематика.
Наприкінці XIX – на початку XX ст. різні пологи жовтозелених відносили до відділу зелених водоростей, що було пов'язано в першу чергу з забарвленням та морфологічною схожістю таломів. В даний час жовтозелені розглядаються як клас усередині відділу охрофітових.
Відомо близько 450 сучасних видів класу Жовтозелених водоростей, які групуються в чотири порядки: Ботрідієві, Мішококкові, Трибонемові та Вошерієві. В основу виділення порядків покладено тип диференціації талому та особливості життєвого циклу.
Порядок Ботридієві – Botrydiales. До порядку відносяться види з сифональним типом диференціації талому, у яких відсутній оогамний статевий процес.
Рід Ботрідіуммешкає на ґрунті і має вигляд зелених пухирів завбільшки кілька міліметрів, прикріплених за допомогою безбарвних ризоїдів. Таллом сифональний, містить численні ядра та пластиди. Оболонка багатошарова, на ній може відкладатися вапно. Розмноження безстатеве за допомогою двожгутикових зооспор, при цьому весь вміст міхура розпадається на одноядерні фрагменти. При нестачі вологи розмножується за допомогою апланоспор або формує товстостінні цисти. В одних випадках весь вміст міхура йде на утворення однієї великої цисти. В інших випадках цисти формуються в ризоидах, куди спочатку переходить вміст міхура. Цисти проростають або у новий таллом, або утворюють зооспори. Статевий процес - ізо-і гетерогамія. Зигота проростає негайно, без періоду спокою. Звичайні та широко поширені в наземних місцеперебуваннях види, що зустрічаються по берегах струмків, ставків або на позбавлених рослинності ґрунтах.
Порядок Місхококові–Mishococcales. Одноклітинні або колоніальні представники з кокоїдним типом диференціації талому.
Рід Хараціопсисвключає прикріплені одноклітинні форми. При розмноженні формує зооспори, апланоспори та товстостінні цисти (рис. 44, 1-2).
Рід Офіоцитіум(Рис. 44, 4) має подовжені циліндричні клітини, які можуть бути прямими, зігнутими або спірально закрученими, а на кінці можуть нести шип. Клітинна стінка і двох нерівних частин, їх більшість бере участь у зростанні клітини, менша частина стала і має форму кришечки. Одноклітинні та колоніальні види, що вільно живуть або прикріплені до субстрату за допомогою маленької ніжки. Розмножуються зооспорами та апланоспорами, зустрічаються цисти. Мешкають у прісних водах.
Рід Місхококкусутворює деревоподібні прикріплені колонії. Розгалуження дихотомічне та тетрахотомічне. Клітини розташовані по 2 чи 4 на верхівках слизових гілок колонії. Клітини кулясті, до овальних, з тонкою або товстою клітинною стінкою. Іноді клітинна стінка блискуча та коричнева через просочення її солями заліза. Молоді одноклітинні організми зі слизовою дископодібною основою, яка служить точкою опори при прикріпленні. Після виходу суперечка протопласт материнської клітини перетворюється на холодець і витягується, довжина стає в 6 разів більшою за ширину, і таким чином виникає циліндрична ніжка. Порожня клітинна стінка материнської клітини завжди стає основою ніжки. Безстатеве розмноження зооспорами та автоспорами. Автоспори прикріплюються до верхнього краю слизової оболонки ніжки. Наступні клітинні поділу повторюють процес і виробляють деревоподібну колонію. Статевий процес – ізогамія. Мешкають у невеликих прісних водоймах як епіфіти ниткових водоростей. Відомі у центральній Європі та Азії.
Порядок Трибонемові – Tribonematales. Представники мають нитчастий, різнонітчастий, ложнотканинний та тканинний тип диференціації талому. Клітинні стінки або з Н-подібними частинами, що перекриваються, або цілісні.
Рід Трибонема– нитки, що не гілкуються (рис. 46, 1). Клітини за формою циліндричні або барилоподібні. Клітинна стінка складається з двох половинок, які знаходять краями одна на одну у середині клітини. Оболонки часто шаруваті. Фрагменти ниток завжди закінчуються порожніми половинками Н-подібних фрагментів оболонки, що мають вигляд вилочки. У клітинах кілька жовтувато-зелених пластид, піреноїдів немає. Розмноження вегетативне (фрагментацією ниток), безстатеве (зооспорами та апланоспорами) та статеве (ізогамія), причому апланоспори утворюються частіше, ніж зооспори. Можуть формувати акінети. Мешкають у прісних водах, де особливо рясно розвиваються в холодну пору року.
Порядок Вошерієві – Vauсheriales. Всі представники мають сифональний талл, оогамний статевий процес та синзооспори.
Рід Вошерія(Рис. 46, 3) має слоевище неклітинної будови; її талом досягає завдовжки кількох сантиметрів, прикріплюється до субстрату за допомогою безбарвного різоїду. У нитках відсутні перегородки, більшість талому зайнята вакуоллю, а, по периферії в цитоплазмі розташовані численні ядра і пластиди. Нитки з апікальним зростанням та рідкісним бічним розгалуженням. Перегородки утворюються при пошкодженні талому та для відділення органів розмноження. Безстатеве розмноження здійснюється апланоспорами, синзооспорами, акінетами. Синзооспори формуються по одній у зооспорангії, що відокремлюється від вегетативних клітин перегородкою на кінці нитки. Зооспори багатоядерні та багатоджгутикові. Статевий процес – оогамія. Зигота покривається товстою оболонкою і після періоду спокою проростає у новий талом.
Види Вошеріїшироко поширені в прісних, солонуватих та морських водах, а також у наземних місцеперебуваннях. Вони зустрічаються всіх континентах, включаючи Антарктиду. Формують трав'яно-зелені або темно-зелені переплутані маси - так звані мати, гладкі, стелиться або подушкоподібні. Водні, напівводні, наземні форми. Мешкають у різноманітних місцеперебуваннях: морях, естуаріях, лиманах, солоних маршах, мангрових чагарниках, струмках, каналах, озерах, ставках, на орних землях і болотах.
Значеннягетероконтних водоростей
З відділу гетероконтних водоростей найбільше значення для природних екосистем і для людини мають бурі водорості.
Бурі водорості – основний джерело органічної речовиниу прибережній зоні морів. Їхня біомаса в морях помірних і приполярних зон може досягати кількох десятків кілограмів на квадратний метр. Зарості бурих водоростей створюють умови для живлення та розмноження безлічі прибережних тварин та інших водоростей. Чарльз Дарвін, який спостерігав біля берегів Південної Америки зарості бурої водорості Макроцистису, писав: «Ці величезні підводні ліси Південної півкулі можу порівняти лише з наземними лісами тропічних областей. І все-таки, якби в якійсь країні знищити ліс, то не думаю, щоб при цьому загинула хоча б приблизно така кількість видів тварин, як із знищенням водоростей».
Зарості бурих водоростей служать місцем харчування,укриття та розмноженнябагатьох тварин. Образно кажучи, бурі водорості надають іншим водним організмам «стіл, дах та ясла».
Широко використовуються бурі водорості та людиною. Вони багаті йодомта іншими мікроелементами. Народи Південно-Східної Азії традиційно використовують їх в їжу, особливо представників порядку ламінарієвих, з яких готують безліч найрізноманітніших страв. Кормове борошно, що готується з бурих водоростей, підвищує продуктивність худоби; при цьому в яйцях та молоці збільшується вміст йоду.
З бурих водоростей отримуютьальгінати- Солі альгінової кислоти. Альгінати дуже широко застосовують у різних галузях промисловості. Це нетоксичні сполуки, що мають колоїдні властивості, тому вони широко використовуються в харчовій та фармацевтичній промисловості. Альгінова кислота та її солі здатні до 200-300-кратного поглинання води, утворюючи гелі, для яких характерна висока кислотостійкість. У харчовій промисловості вони використовуються переважно як емульгатори, стабілізатори, желюючі та вологоутримуючі компоненти. Наприклад, сухий порошковий альгінат натрію використовують у виробництві порошкоподібних та брикетованих розчинних продуктів (кава, чай, сухе молоко, киселі та ін) для їх швидкого розчинення. Водні розчини альгінатів використовують для заморожування м'ясних та рибних продуктів. У світі харчову промисловість йде до 30% всього обсягу одержуваних альгінатів.
У текстильній та целюлозно-паперовій промисловості альгінати використовують для загущення фарб та посилення міцності їх зв'язку з основою. Просочення тканин деякими альгінатами надає їм захисних властивостей: водонепроникність, кислотостійкість і збільшує механічну міцність. Ряд солей альгінових кислот використовують для отримання штучного шовку. За часів Другої світової війни в США та Англії з альгінової кислоти та її солей вироблялася велика кількість маскувальної тканини та мереж для житлових та промислових будівель.
Альгінати застосовують у металургії: у ливарному виробництві вони покращують якість формувальної землі. Солі альгінової кислоти застосовують у виробництві електродів для електрозварювання, що дозволяють отримувати більш якісні зварні шви. Альгінати також використовують при виробництві пластмас, синтетичних волокон, лакофарбових покриттів та будівельних матеріалів, стійких до атмосферних впливів. Їх застосовують при виготовленні високоякісних мастильних матеріалів для машин. У радіоелектроніці альгінати виконують роль сполучного агента для виготовлення високоякісних феритів.
Найбільше застосування знаходить водорозчинний альгінат натрію, здатний до утворення в'язких розчинів. Його широко використовують для стабілізації різноманітних розчинів та суспензій. Додавання невеликої кількості альгінату натрію в харчові продукти – консерви, морозиво – підвищує їхню якість. Він також використовується для виготовлення декоративної косметики, кремів та масок у парфумерній промисловості.
У фармацевтичній промисловості альгінати використовуються для покриття таблеток, пігулок, як компонентні основи для різних мазей і паст, як гелі-носії лікарських препаратів, у виробництві розчинних хірургічних ниток. У медицині альгінат кальцію застосовують як кровоспинний засіб і як сорбент, що виводить радіонукліди (наприклад, стронцій). Щорічне виробництво альгінатів у світі перевищує 20 тисяч тонн.
Інша важлива речовина, що отримується з бурих водоростей, – шестиатомний спирт маніт. Маніт використовують як замінник цукру для хворих на діабет. Крім того, він може бути використаний як плазмозамінник при консервації крові. Його застосовують для виготовлення таблеток у фармацевтичній промисловості. Маніт також використовують у виробництві синтетичних смол, фарб, паперу, вибухових речовин при виробленні шкір.
Фукоїдани,одержувані з бурих водоростей – ефективні антикоагулянти, навіть активніші, ніж гепарин. Перспективним вважається використання їх для отримання протипухлинних препаратів та антивірусних сполук. Адже навіть у найнижчих концентраціях фукоїдани можуть інгібувати прикріплення вірусів до поверхні клітин. Фукоїдани також мають здатність утворювати виключно міцні і в'язкі слизи, що знаходить застосування в отриманні стабільних емульсій та суспензій.
Енергетична криза, що охопила в останні роки багато країн світу, призвела до необхідності пошуку нових нетрадиційних джерел енергії. Так, у США з цією метою вивчається можливість розведення гігантської ламінарієвої водорості Макроцистисз подальшою переробкою її в метан. Підраховано, що з площі 400 км2, зайнятої цією водорістю, можна отримати 620 млн. м3 метану.
Гетероконтні водорості із класів Золотисті, Жовтозелені, Синурові, Рафідофітові та Євстигмові водорості представлені, в основному, мікроскопічними організмами, широко поширені в прісних водоймах усіх кліматичних зон земної кулі, але частіше зустрічаються в помірних широтах. Є серед золотистих водоростей види, що мешкають у морях та солоних озерах, а дуже мало хто – у забруднених водах. Максимуму розвитку золотисті водорості досягають у холодну пору року: у планктоні вони домінують ранньою весною, пізно восени та взимку. У цей час вони відіграють значну роль як продуценти первинної продукції та служать їжею організмам зоопланктону.
Деякі золотаві водорості, наприклад Урогленаі Динобріон,розвиваючись у масовій кількості, здатні викликати цвітіння води. Вони виділяють альдегіди та кетони, які можуть надавати воді неприємного запаху та смаку. Уроглена– токсичні для риб жирні кислоти
Рафідові водорості широко представлені в планктоні прісних водойм переважно з кислим рН, особливо в сфагнових болотах, рідше у великих озерах. У прісних водоймах локальні «цвітіння» може формувати Гоніостомум.Рафідові водорості зустрічаються також у опріснених морських бухтах та калюжах на березі моря, а також у відкритому морі. При масовому розвитку у прибережних морських водах вони викликають токсичне "цвітіння" води. Так, біля узбережжя Канади під час "цвітіння" концентрація клітин рафідової водорості Гетеросигми може досягати 30 млн. на 1 л. Спалахи чисельності рафідових водоростей часто призводять до розвитку «червоних припливів», з якими пов'язують загибель риб. Причиною таких «червоних припливів» можуть бути види пологів Хаттонелла, Олістодіскус, Гетеросигма та Фіброкапса.
Синурові водорості при масовому розвитку в прісних водоймах можуть надавати воді неприємного запаху ( Синура). Феотамнієві водорості зустрічаються у стоячих та повільно поточних прісних водоймах, де поселяються епіфітно на ниткових водоростях.
Євстигмові водорості зустрічаються лише у прісних водоймах чи грунті.
Жовтозелені водорості поширені на всіх континентах, вони мешкають переважно в прісних водах і ґрунті, а також у наземних, солонуватоводних та морських місцеперебуваннях. Жовтозелені водорості населяють чисті та забруднені води, з різним значенням рН: можуть мешкати як у кислих, так і в лужних водах. Вони зустрічаються, головним чином, у чистих прісноводних водоймищах, рідше в морях і солонуватих водах, віддаючи перевагу помірній температурі, частіше розвиваються навесні і восени, хоча є види, що зустрічаються протягом усіх періодів року, у тому числі і взимку. Найчастіше їх можна знайти в скупченнях нитчаток і серед чагарників вищих водних рослин у прибережній зоні річок, ставків, озер та водосховищ.
Переважна більшість жовтозелених – вільноживучі форми, але зустрічаються і внутрішньоклітинні симбіонти – зооксантели у найпростіших клітинах. Цікавий внутрішньоклітинний симбіоз утворюють хлоропласти морського виду Вошеріїз молюском Елізія. Упродовж дев'яти місяців цей молюск здатний у культурі до фотоавтотрофної фіксації вуглекислого газу. Це найтриваліший у часі симбіоз такого типу, коли симбіотична пластида знаходиться у безпосередньому контакті з цитоплазмою тварини. У природі личинки молюска живляться нитками Вошерії. Внаслідок фагоцитозу хлоропласти водорості потрапляють у цитоплазму епітеліальних клітин молюска. В ході цього процесу оболонка хлоропласту стає тришаровою, втрачається одна зовнішня мембрана ХЕС (хлоропластної ендоплазматичної мережі). Подібне явище є добрим доказом того, що в ході еволюції в результаті вторинного симбіогенезу за рахунок втрати мембран могли виникати хлоропласти з трьома мембранами.
Жовтозелені, золотисті та інші гетероконтні водорості є продуцентами кисню та органічних речовин; вони входять до складу харчових кіл. Гетероконтофіти беруть участь у самоочищенні забруднених вод та ґрунтів, утворенні сапропелю, у процесі накопичення органічних речовин у ґрунті, впливаючи на його родючість. Вони використовують як показові організми щодо стану забруднення вод; жовто-зелені водорості входять до складу комплексу мікроорганізмів, що застосовується для очищення стічних вод.
Контрольні питання
Назвіть характерні риси будови бурих водоростей.
Особливості будови таломів бурих водоростей.
Як розмножуються бурі водорості? Що таке моноспори, тетраспори та зооспори, ізогамія, гетерогамія та оогамія?
Які життєві цикли у бурих водоростей?
Розмноження фукусових та ламінарієвих водоростей.
Назвіть характерні риси та типових представників порядків бурих водоростей.
У яких місцепроживання зустрічаються бурі водорості? Яке значення їх у природі.
Господарське значення бурих водоростей.
Назвіть характерні риси будови та типових представників золотистих водоростей.
Які пігменти та типи харчування відомі у золотистих водоростей?
Розмноження та екологія золотистих водоростей.
Назвіть характерні риси та типових представників жовтозелених водоростей.
Які пігменти та типи харчування відомі у жовтозелених водоростей?
Як розмножуються жовтозелені? Види статевого розмноження: ізогамія, гетерогамія та оогамія?
Назвіть характерні риси будови та типових представників синурових водоростей.
Розмноження та екологія синурових водоростей.
Назвіть характерні риси будови та типових представників феотамнієвих водоростей.
Розмноження та екологія феотамнієвих водоростей.
Назвіть загальні риси будови та типових представників рафідофітових водоростей.
Розмноження та екологія рафідофітових водоростей.
Назвіть загальні риси будови та типових представників евстигмових водоростей.
Розмноження та екологія евстигмових водоростей.
Значення гетероконтних водоростей у природних екосистемах. Одноклітинні, цінобіальні та колоніальні форми водоростей з монадною (джгутиковою) структурою тіла; до них відносяться майже всі Euglenophyta, б.год.
- Pyrrophyta та Chrysophyta, окремі кл. Xanthophуta та Chlorophyta.
- див. Н-антигени...
- - Група нижчих, чаші водних р-ній. Одноклітинні або багатоклітинні, одиночні та колоніальні форми величиною від дек. мікрометрів до десятків метрів. У багатоклітинних Ст слоевище часто розгалужене,...
Сільсько-господарський енциклопедичний словник
- - нижчі рослинні організми. Тіло Ст відрізняється простим будовою і не розчленовується, як у вищих квіткових рослин, на корінь, стебло і листя.
Сільськогосподарський словник-довідник
- - Група переважно. вод. організмів, що зазвичай містять хлорофіл і виробляють органічні. в-ва в процесі фотосинтезу.
Природознавство. Енциклопедичний словник
- - група нижчих верствових рослин, що мають хлорофіл і здатні до фотосинтезу. Розділяються на 6 типів: різножгутикові, бурі, червоні, зелені, діатомові, синьо-зелені.
Тлумачний словник з ґрунтознавства
- - клас найпростіших. Одноклітинні організми, які мають на певній стадії розвитку хлистоподібними пристосуваннями для пересування. Джгутики ж повідомляють їм чутливість.
Науково-технічний енциклопедичний словник
- - велика і неоднорідна група примітивних, що нагадують рослини організмів.
Енциклопедія Кольєра
- - одноклітинні, цінобіальні та колоніальні форми водоростей із монадною структурою тіла; до них відносяться майже всі Euglenophyta, б. ч. Pyrrophyta та Chrysophyta, окремі кл. Xanthophуta та Chlorophyta...
Словник ботанічних термінів
- - див. ризопласти...
Словник ботанічних термінів
Великий медичний словник
- - мікроскопічні одноклітинні, одноджгутикові автотрофні морські планктонні водорості, що мають внутрішньоклітинний кремнеземний, зазвичай порожнистий скелет.
Геологічна енциклопедія
- - частини канальцевої системи губок, вистелені клітинами зі джгутом кожна, оточеним протоплазматичним коміром.
Енциклопедичний словник Брокгауза та Євфрона
- - биченосці, одноклітинні та колоніальні організми, що мають джгутики як органи руху. Деякі групи Ж., наприклад евгленових, ботаніки відносять до рослин, а зоологи - до тварин.
Велика Радянська Енциклопедія
- - Мн., Р.
Орфографічний словник російської мови
- - джгутикові мн. Клас одноклітинних типу найпростіших, що характеризуються наявністю у них джгутиків.
Тлумачний словник Єфремової
- - палив"...
Російський орфографічний словник
"ЖГУТИКОВІ ВОДОРОСЛИ" у книгах
автора З книги Тваринний світ Дагестану автора Шахмарданов Зіяудін Абдулганієвіч1.2. Тип Саркомастигофори, або Саршкіргутиконосці, Саркодові та Жгутикові (Sarcomastigophora)
З книги Тваринний світ Дагестану автора Шахмарданов Зіяудін Абдулганієвіч1.2. Тип Саркомастигофори, або Саршкіргутиконосці, Саркодові та Жгутикові (Sarcomastigophora) Їх 18000 видів, мають ложноніжки - непостійні вирости цитоплазми або джгутики, а іноді й ті, й інші. Тип має два класи: 1) Саркодові
1.2.2. Клас Жгутиконосці (Mastigophora), або Жгутикові, Біченосці
З книги Тваринний світ Дагестану автора Шахмарданов Зіяудін Абдулганієвіч1.2.2. Клас Жгутиконосці (Mastigophora), або Жгутикові, Біченосці До джгутиконосців (джгутикові) відносяться понад 6 тис. видів найпростіших, одноклітинні та колоніальні організми, що мешкають у морях, прісних водоймах, у ґрунті та в організмах багатоклітинних тварин. Форма тіла їх
Водорості
З книги Суші та роли готуємо вдома автора Костіна ДарьяВодорості
автора Волкова ТрояндаБурі водорості
З книги Дари моря, які зцілюють організм автора Волкова ТрояндаВодорості
З книги Дари моря, які зцілюють організм автора Волкова ТрояндаВодорості Нижчі рослини У ботаніків прийнято ділити всі рослини на вищі та нижчі. У повсякденному житті ми з вами зустрічаємося в основному з вищими рослинами, тобто з тими, які мають листя, стебла і коріння. До вищих рослин відносяться мохи, плауни, хвощі та папороті,
Водорості
З книги Довідник кросвордиста автора Колосова СвітланаВодорості
З книги Історія нашого обману, або Як харчуватися, чим лікуватися, як не опромінитися, щоб залишитися здоровим автора Мізун Юрій ГавриловичВодорості До складу водоростей входять практично всі вітаміни та найважливіші мікро- та макроелементи, наприклад йод. Він розщеплює жирові елементи, які потім легко виводяться з організму. Водорості також виводять шлаки і зміцнюють шкіру. У багатьох морських водоростях
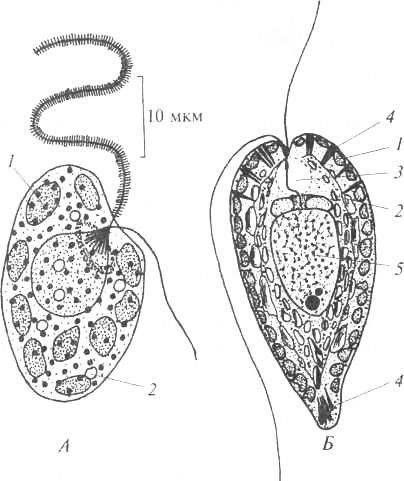
Монадні вегетативні клітини та монадні стадії у життєвому циклі (зооспори та гамети) водоростей забезпечені джгутиками – довгими та досить товстими виростами клітин, зовні покритими плазмалемою. Їхня кількість, довжина, морфологія, місце прикріплення, характер руху досить різноманітні у водоростей, але постійні всередині родинних груп.
Джгутики можуть прикріплюватися на передньому кінці клітини (апікально) або можуть бути трохи зрушені вбік (субапікально); можливе їх прикріплення збоку клітини (латерально) та на черевній стороні клітини (вентрально). Джгутики, однакові за морфологією, називають ізоморфними, якщо вони різняться гетероморфними. Ізоконні- це джгутики однакової довжини, гетероконтні- Різної довжини.
Джгутики мають єдиний план будівлі. Можна виділити вільну частину (ундуліподію), перехідну зону, базальне тіло (кінетосому). Різні частини джгутика відрізняються за кількістю та розташуванням мікротрубочок, які утворюють скелет (рис. 2).
Мал. 2. Схема будови джгутиків водоростей (по: Л.Л.Великанов та ін., 1981): 1 – поздовжній розріз джгутиків; 2, 3 – поперечний розріз через кінчик джгутика; 4 – поперечний розріз через ундуліподію; 5 – перехідна зона; 6 – поперечний розріз через основу джгутика – кінетосому
Ундуліподія(у перекладі з латинського «хвильовика») здатна здійснювати ритмічні хвилеподібні рухи. Ундуліподія є одягненою мембраною аксонему. Аксонемаскладається з дев'яти пар мікротрубочок, розташованих по колу, і пари мікротрубочок у центрі (рис. 2). Джгутики можуть бути гладкими або вкриті лусочками або мастигонемами (волосками), а у динофітових і криптофітових вони покриті лусочками та волосками. Джгутики примнезіофітових, криптофітових та зелених водоростей можуть бути покриті лусочками різної форми та розмірів.
Перехідна зона. Функціонально відіграє роль у зміцненні джгутика у місці його виходу з клітини. У водоростей виділяють кілька типів структур перехідної зони: поперечна пластинка (дінофітові), зірчаста структура (зелені), перехідна спіраль (гетероконтні), перехідний циліндр (примнезіофітові та дінофітові).
Базальне тіло чи кінетосома. У цій частині джгутика структура у вигляді порожнистого циліндра, стінка якого утворена дев'ятьма триплетами мікротрубочок. Функція кінетосоми – зв'язок джгутика із плазмалемою клітини. Базальні тіла в ряду водоростей можуть брати участь у розподілі ядра і ставати центрами організації мікротрубочок.
Кінець роботи -
Ця тема належить розділу:
Водорості
Рибогосподарський університет.. інститут біології моря ім у жирмунського два ран.. л л кавуна..
Якщо Вам потрібний додатковий матеріал на цю тему, або Ви не знайшли те, що шукали, рекомендуємо скористатися пошуком по нашій базі робіт:
Що робитимемо з отриманим матеріалом:
Якщо цей матеріал виявився корисним для Вас, Ви можете зберегти його на свою сторінку в соціальних мережах:
| Твітнути |
Всі теми цього розділу:
Клітинні покриви
Клітинні покриви забезпечують стійкість внутрішнього вмісту клітин до зовнішніх впливів і надають клітин певну форму. Покриви проникні для води та розчинених у ній низькомолекул.
Мітохондрії
Мітохондрії зустрічаються в клітинах еукаріотних водоростей. Форма та будова мітохондрій у клітинах водоростей різноманітніші порівняно з мітохондріями вищих рослин. Вони можуть бути округлими
Пігменти
Всі водорості добре розрізняються за набором фотосинтезуючих пігментів. Такі групи у систематиці рослин мають статус відділів.
Основним пігментом усіх водоростей є зелений пігмен
Пластиди
Пігменти в клітинах еукаріотних водоростей знаходяться у пластидах, як і у всіх рослин. У водоростей зустрічається два типи пластид: пофарбовані хлоропласти (хроматофори) та безбарвні лейкопласти (амі
Ядро та мітотичний апарат
Ядро водоростей має типову для еукаріот структуру. Кількість ядер у клітині може варіювати від одного до кількох. Зовні ядро покрите оболонкою, що складається з двох мембран, зовнішня мемб
Монадний (джгутиковий) тип структури талому
Найбільш характерною ознакою, що визначає цей тип структури, є наявність джгутиків, за допомогою яких монадні організми активно пересуваються у водному середовищі (рис. 9, А). Рухливі ж
Ризоподіальний (амебоїдний) тип структури
Найбільш суттєві ознаки амебоїдого типу структури - відсутність міцних клітинних покривів і здатність до амебоїдного руху, за допомогою ци, що тимчасово утворюються на поверхні клітини ци
Пальмеллоїдний (гемімонадний) тип структури
Характерним для цього типу структури є поєднання нерухомого рослинного способу життя з наявністю клітинних органел, властивих монадним організмам: скорочувальні вакуолі, стигма, джгут
Кокоїдний тип структури
Цей тип поєднує одноклітинні та колоніальні водорості, нерухомі у вегетативному стані. Клітини кокоїдного типу одягнені оболонкою і мають протопласт рослинного типу (тонопласт без сократ
Трихальний (нитчастий) тип структури
Характерною особливістю нитчастого типу структури є ниткоподібне розташування нерухомих клітин, які утворюються вегетативним шляхом в результаті клітинного поділу, що відбувається переважно.
Гетеротрихальний (різнонитчастий) тип структури
Різнонитчастий тип виник на основі нитчастого типу. Різнонітчаста слоевище складається здебільшого з горизонтальних, що стелиться по субстрату ниток, що виконують функцію прикріплення, і вертикальних,
Паренхіматозний (тканинний) тип структури
Один із напрямів еволюції різнонітчастої слані було пов'язане з виникненням паренхіматозних слоев. Здатність до необмеженого зростання та поділу клітин у різних напрямках призвела до обра
Сифональний тип структури
Сифонокладальний тип структури
Основною ознакою сифонокладального типу структури є здатність до утворення з первинного неклітинного слані складно влаштованих сланів, що складаються з первинно багатоядерних сегментів. У
Безстатеве розмноження
Безстатеве розмноження водоростей здійснюється за допомогою спеціалізованих клітин - суперечка. Спороутворення зазвичай супроводжується розподілом протопласту на частини та виходом продуктів поділу з
Просте розподіл
Цей спосіб розмноження зустрічається лише в одноклітинних форм водоростей. Найбільш просто відбувається розподіл у клітин, що мають амебоїдний тип будови тіла.
Розподіл амебоїдних форм
Фрагментація
Фрагментація властива всім групам багатоклітинних водоростей і проявляється в різних формах: утворення гормогоніїв, регенерація частин слані, що відірвалися, спонтанне відпадання гілок, відростання
Розмноження пагонами, столонами, виводковими бруньками, бульбочками, акінетами
У тканинних форм зелених, бурих та червоних водоростей вегетативне розмноження набуває своєї закінченої форми, яка мало відрізняється від вегетативного розмноження вищих рослин. Зберігаючи спосо
Статеве розмноження
Статеве розмноження у водоростей пов'язане зі статевим процесом, який полягає у злитті двох клітин, внаслідок чого утворюється зигота, що виростає в нову особину або дає зооспори.
Зміна ядерних фаз
При статевому процесі внаслідок злиття гамет та їх ядер відбувається подвоєння числа хромосом у ядрі. На певному етапі циклу розвитку, при мейозі, відбувається редукція числа хромосом, в результаті
Ендофити/ендозоїти, або ендосімбіонти
Ендосимбіонти, або внутрішньоклітинні симбіонти - водорості, що живуть у тканинах або клітинах інших організмів (безхребетних тварин або водоростей). Вони утворюють своєрідну екологічну гру
Відділ синьо-зелених водоростей (ціанобактерії) – cyanophyta
У назві відділу (від грец. cyanos – синій) відбито характерну особливість цих водоростей – забарвлення талому, пов'язане з відносно високим вмістом синього пігменту фікоціаніну. Ціан
Порядок – Chroococcales
Зустрічаються як одноклітинних " простих " індивідів чи частіше утворюють слизові колонії. При розподілі клітин у двох площинах виникають одношарові пластинчасті колонії. Розподіл у трьох п
Відділ червоних водоростей- rhodophyta
Назва відділу походить від грецького слова rhodon ("родон") -рожевий. Забарвлення червоних водоростей обумовлене різним поєднанням пігментів. Вона буває від сірого та фіолетового
Рід Порфіра має слоевище у вигляді тонкої блискучої пластини з рівними або складчастими краями, що складається з одного-двох шарів щільно з'єднаних клітин. Основа пластини зазвичай переходить у
Порядок Родименіеві – Rhodymeniales
Рід Спарлінгія (Батьківщина) – плоскі пластини до 45 см висоти, листоподібної та клиноподібної форми, розширені та пальчасто розсічені вгорі, від світло-рожевого або світло-оранжевого до
Порядок Коралінові – Corallinales
Рід Коралліна - членисті віялоподібно розгалужені кущики до 10 см висоти, розгалужені, вапняні, від рожево-бузкового до майже білого кольору. Розмножується безстатевим та статевим шляхом. Спо
Порядок Гігартинові – Gigartinales
Рід Хондрус - щільні шкірясті хрящуваті кущики до 20 см висоти, 3-4-разово розгалужені, світло-жовтого, світло-рожевого, пурпурно-темно-червоного кольору. Зростає в нижній частині літоралі та
Порядок Церамієві – Ceramiales
Рід Цераміум - ніжні пухнасті членисті кущики до 10 см висоти, дихотомічно або по черзі розгалужені, темно-жовтого з рожевим відтінком кольору. Розгалуження двох-чотирьох порядків, кінець
Відділ діатомових водоростей – bacillariophyta
Відділ називається Діатомові водорості (від грец. di – два, tome – розріз, розтин), або Бацилярії (bacillum – паличка). Включає одноклітинні одиночні або колоніальні орг
Відділ гетероконтні (різножгутикові) водорості – heterokontophyta
У всіх гетероконтів схоже влаштований джгутиковий апарат. Є 2 джгутики, причому на одному з них є дуже характерні трубчасті тричленові перисті вирости, або волоски - мастигонеми. Саме готівка
Систематика
Викопні коколіти відомі з мезозойських відкладень і були рясні протягом більшої частини юрського та в крейдяному періодах. Максимального розмаїття примнезіофітові досягли в пізньому крейді,
Відділ криптофітових водоростей (криптомонади) - cryptophyta
Відділ названий за типовим родом криптомонас (від грец. kryptos - прихований, monas - особина). Включає одноклітинні, рухливі, монадні організми. Клітини криптофітів
А Б В Г Д
Мал. 53. Зовнішній вигляд криптофітових водоростей (по: Г.А. Белякова та ін., 2006): А - Родомонас, Б - Хроомонас, В - Криптомонас, Г - Хіломонас, Д - Гоніомонас може фор
Відділ зелених водоростей – chlorophyta
Зелені водорості - найбільший із усіх відділів водоростей, що налічує за різними оцінками від 4 до 13 - 20 тисяч видів. Всі вони мають зелений колір слані, що обумовлено переважанням в хлоро
Порядок Улотриксові – Ulotrichales
Рід Улотрікс (рис. 54). Види Улотрикса мешкають частіше в прісних, рідше в морських, солонуватих водоймах і грунті. Вони прикріплюються до підводних предметів, формуючи яскраво-зелені кущики.
Порядок Бріопсидові-Bryopsidales
Більшість видів зустрічається в прісних та солонуватих водоймах. Деякі з них ростуть на ґрунті, на камінні, піску та іноді на солончаках.
Рід Бріопсис – ниткоподібні кущики до 6-8 с
Порядок Вольвоксові - Volvocales
Рід Хламідомонада (рис. 57) включає понад 500 видів одноклітинних водоростей, які мешкають у прісних, дрібних, добре прогріваються і забруднених водоймищах: ставках, калюжах, канавах і т.п. Пр
Відділ Харофітові (Харові) – Charophyta
Харофітові – лінія прісноводних зелених водоростей, що призвела до вищих рослин. Це форми переважно з нитчастим таломом. Нерідко талом вертикальний, розчленований і несе про
Відділ динофітові (динофлагеллати) - dinophyta
1. Назва відділу походить від грецьк. dineo – обертатися. Об'єднує переважно одноклітинні монадні, рідше кокоїдні, амебоїдні або пальмелоїдні, іноді колоніальні.
Відділ euglenozoa – євгленові
Відділ назва за типовим родом - Euglena (від грец. еu – добре розвинений, glene – зіниця, око). Поєднує одиночних монадних чи амебоїдних представників. Зрідка зустрічаються до
Словник термінів
У всіх гетероконтів схоже влаштований джгутиковий апарат. Є 2 джгутики, причому на одному з них є дуже характерні трубчасті тричленові перисті вирости, або волоски - мастигонеми. Саме наявність двох нерівних джгутиків стало підставою для того, щоб назвати цю велику групу водних організмів. різножгутиковимиводоростями, або гетероконтами. Крім відмінностей у довжині, джгутики розрізняються і морфологічно: головний джгутик складається з осі та перисто розташованих на ній миготливих волосків, бічний джгутик гладкий бичевидний. Подібний також склад пігментів – хлорофіли аі зта численні каротиноїди, а також будова хлоропластів. Хлоропласти оточені чотирма мембранами, причому дві зовнішні мембрани пов'язані з ендоплазматичною мережею клітини.
У складі відділу Гетероконтні (Різножгутикові) водорості поєднують кілька груп водоростей у ранзі класів: Бурі, Золотисті, Жовтозелені., Рафідофітові, Синурові, Феотамнієві, Євстигмові, деякі раніше розглядалися як самостійні відділи.
Клас Бурі водорості– Phaeophyceae (Fucophyceae)
Назва класу «бурі водорості» відображає забарвлення їхнього талому (від грец. рhaios- Смаглявий). Забарвлення обумовлене наявністю великої кількості бурих та жовтих пігментів – крім хлорофілів аАвтогамія – статеве розмноження, при якому зливаються два сестринські гаплоїдні ядра в загальній цитоплазмі. зАвтогамія – статеве розмноження, при якому зливаються два сестринські гаплоїдні ядра в загальній цитоплазмі. β -каротину в їх хлоропластах є надлишок ксантофілів, особливо фукоксантину.
Багатоклітинні, переважно великі морські водорості, особливо поширені в холодних водах обох півкуль. Ні одноклітинні, ні колоніальні форми серед бурих водоростей невідомі. Відомо близько 1,8 тис. видів.
Будова клітини. Клітина у видів бурих водоростей еукаріотна: вона має оформлені органели з власними мембранами: ядро, мітохондрії, пластиди та ін. Зелені пігменти – хлорофіли аі з– маскуються додатковими: помаранчевими каротинами та жовто-коричневими ксантофілами; з останніх переважає фукоксантін. У клітинах бурих водоростей зустрічається від однієї до багатьох пластид. Поверх зовнішньої клітинної мембрани клітини бурих водоростей зазвичай виробляють тверду клітинну стінку.
Ядро найчастіше одне, але іноді ядер кілька. У деяких представників із збільшенням віку клітини кількість ядер збільшується. Ядро в ядрі завжди одне.
Хлоропласти вегетативних клітин бурих водоростей мають вічко (стигму) та піреноїди, яких буває один або кілька; у деяких видів піреноїди відсутні. Вічко, що складається з 30-40 ліпідних глобул, виконує функцію лінзи, що фокусує світло на джгутикове здуття, яке є фоторецептором. Зооспори та гамети бурих водоростей зазвичай не мають вічка. У піреноїдах відкладаються запасні речовини. Хлоропласти найчастіше дрібні, дископодібні, парієтальні. Їх форма може бути зірчастою, стрічковоподібною або пластинчастою. Форма хлоропластів у клітинах бурих водоростей може змінюватись у міру старіння клітини. Оболонка хлоропласту складається з чотирьох мембран, причому дві зовнішні мембрани пов'язані з ЕПС-ендоплазматичною мережею клітини.
Целюлоза, з якої складається стінка клітин бурих водоростей, її іноді називають альгулозою, за властивостями відрізняється від целюлози вищих рослин. Назовні від целюлозної стінки розташовується пектиновий шар, що сильно ослизняється, утворений в основному альгінової кислотою і її солями і сполуками з білковими речовинами. Альгінова кислотатакож виявлена лише у бурих водоростей.
Автоспора – структура безстатевого розмноження, що є
Всі бурі водорості є фототрофамиі хризоламінарин,спирт маніті ліпіди.Ліпіди накопичуються у вигляді крапель олії. Хризоламінарин відкладається за межами хлоропласту: він розташовується на спеціальних вакуолях на цитоплазмі у піреноїду. Маніт виконує резервну, а також осморегуляторну функцію; його концентрація усередині клітини залежить від зміни солоності води.
У молодих клітинах бурих водоростей містяться дрібні та численні вакуолі, які з віком стають більшими за рахунок злиття. У цитоплазмі розташовані особливі освіти – фізиди, які містять флоротаніни.Ці речовини відомі лише у видів бурих водоростей. Функція флоротанінів, можливо, пов'язана із захистом таломів водоростей від виїдання тваринами, наприклад, молюсками. Флоротаніни бурих водоростей, ймовірно, інгібують поселення на поверхні талому епіфітних водоростей та тварин. Передбачається, що флоротаніни беруть участь у захисті від радіційного пошкодження та у виборчому поглинанні з водного середовища іонів важких металів. Безбарвні флоротаніни на повітрі окислюються з утворенням пігменту фікофеїну, який надає висушеним бурим водоростям характерного темного забарвлення.
Будова талому.Серед видів бурих водоростей відсутні одноклітинні та колоніальні форми, всі особини багатоклітинні. Форма таломів бурих водоростей найрізноманітніша. Це нитки, що стелиться або вертикально стоять, однорядні або багаторядні; масивні суцільні або черепітчасто з'єднані кірки та скоринки; тонкі слизові пластинки або щільні шкірясті пластини на стовбурі, прості або розсічені; бульбашки та мішки; трубки та шнури; по-різному розгалужені тонкі кущі або потужні кущі. Прикріплення таломів здійснюється за допомогою ризоїдів або підошви. Для утримання у вертикальному положенні у великих бентосних форм бурих водоростей утворюються повітряні (плавальні) бульбашки, заповнені газом (рис. 29).
Мал. 29. Зовнішній вигляд бурих водоростей: 1 – Фукус; 2 – Сахаріна; 3 – Аларія; 4 – Саргассум
Найбільш складно серед видів бурих водоростей влаштовані таломи представників порядків Ламінарієві та Фукусові. Вони мають ознаки тканинного диференціювання зі спеціалізацією клітин талому (рис. 30). У їх слоевищах можна розрізнити кору, що складається з інтенсивно пофарбованих клітин та серцевину, Що складається з безбарвних клітин, які часто зібрані в нитки. У Ламінарієвих у серцевині утворюються також ситоподібні трубки та трубчасті нитки. Серцевина виконує як транспортну функцію, а й механічну, оскільки у ній перебувають нитки з товстими поздовжніми стінками. Між корою і серцевиною у багатьох бурих водоростей може бути проміжний шар великих безбарвних клітин.
Зрістталому у представників бурих водоростей найчастіше інтеркалярний (вставковий) та апікальний (верхівковий), рідше базальний. Інтеркалярне зростання може бути дифузним, або є зона зростання. У великих форм вставкова (інтеркалярна) меристема, або група клітин, що активно ділиться, розташована в місці переходу «черешка» в «листову» платівку. Великі водорості також мають меристематичну зону на поверхні талому, так звану меристодерму (своєрідний аналог камбію вищих рослин).
Розмноженняу бурих водоростей вегетативне, безстатеве та статеве. Гамети формуються в багатогніздних гаметангіях, зооспори - в 
Мал. 30. Внутрішня будова Ламінарії(по: Л.Л. Великанов та ін., 1991): А- Поздовжній зріз стволика; БЗагальна характеристика. У– зріз пластини із сорусами зооспорангіїв
одногніздних спорангіях.
Вегетативне розмноження у ряду видів бурих водоростей може здійснюватися ділянками талому; у видів роду Сфацелярія– вивідковими гілочками (висновковими нирками); у видів роду Фукус- групою клітин на підошві, які здатні до диференціювання у новий талом.
Безстатеве розмноження бурих водоростей зазвичай відбувається за допомогою зооспор. Деякі види розмножуються нерухомими тетраспор або моноспор. Спори безстатевого розмноження формуються в результаті мейозу і наступних мітозів одногніздних спорангіях. Статевий процес – ізогамія, гетерогамія та оогамія. Гамети утворюються в багатогнездних гаметангіях- Спеціальних групах клітин. У кожному гнізді гаметангія формується одна гамета.
Для бурих водоростей відомі статеві феромони- Розчинні речовини, які координують активність клітин при розмноженні. Феромони, яких відомо щонайменше десять, можуть або стимулювати розкриття антеридіїв, або залучати чоловічі гамети до жіночих. Вони також беруть участь у ізоляції видів. Вважають, що феромони утворюються у більшості видів бурих водоростей, що мають статевий процес.
Життєвий цикл.У бурих водоростей можна назвати два основних типи життєвих циклів. Один тип - гаплодіплобіонтний. Спорофити диплоїдні, тобто. усі клітини містять повний набір хромосом. Суперечки безстатевого розмноження формуються в одногніздних спорангіях. При утворенні суперечка відбувається редукційний поділ – мейоз. Така редукція називається споричною. Клітини (спори), що утворилися після мейозу, гаплоїдні, тобто. містять половинний набір хромосом. Гаплоїдні зооспори та тетраспори проростають у гаплоїдний гаметофіт, на якому у багатогніздних гаметангіях формуються гамети. Диплоїдна зигота, що утворилася після злиття гамет, проростає в диплоїдний спорофіт.
Інший тип – диплобіонтний; спорофіти, суперечки та гаметофіти диплоїдні; редукційний поділ відбувається при утворенні гамет. Така редукція називається гаметичною. Після злиття гаплоїдних гамет утворюється диплоїдна зигота і проростає в диплоїдний спорофіт. Цей тип поширений у видів ламінарієвих (рис. 31).

Мал. 31. Життєвий цикл Сахаріни(по: Л.Л.Великанов та інших., 1991): 1 – спорофит, 2 – сорусы зооспорангиев, 3 – зооспори , 4 – гаметофіт, 5 – антеридій, 6 – оогоній, 7 – антерозоїд, 8 –зигота (запліднена яйцеклітина)
На макроскопічному спорофіті Сахаріни, у верхній частині пластини, смугами розвиваються соруси(групи) зооспорангіїв. У зооспорангіях після редукційного поділу утворюються зооспори з двома нерівними джгутиками. Вони виростають у мікроскопічні гаметофіти: чоловічі з антеридіями та жіночі з оогоніями. Запліднена антерозоїдам яйцеклітина відразу, без періоду спокою, проростає і утворює спорофіт. На цьому життєвий цикл Сахарінизавершується.
Фукусові водорості немає спорофиов. У Фукусчоловічі та жіночі статеві органи – антеридії зі сперматозоїдами та концептакули з оогоніями та яйцеклітинами – знаходяться у поглибленнях слані – скафідіях, чоловічі та жіночі скафідії можуть перебувати на одному рецептакуліабо у різних рецептакулах. Оогонії сидять на клітці-ніжці , що виникає на стінці концептакула(Рис. 32, В, Р).Клітка оогонія зазнає трьох поділів

Мал. 32. Розмноження Фукус(по: B. Fott, 1971; W. Braune et al., 1974): A - Фрагмент талому (1 - повітряний міхур, 2 - Рецептакул); Б- Розріз через рецептакул; У– розріз жіночої скафідії; Г– оогоній; Д- Вихід яйцеклітин; Е- Розріз чоловічого скафідія; Ж– гілочка з антеридіями; 3 – вихід сперматозоїдів
ядра, перше з яких редукційне, у результаті в оогонії утворюється 8 яйцеклітин.
Антеридії зазвичай формуються на підтримують їх розгалужених нитках – парафізах(Рис. 32, Е,Ж).В результаті декількох поділів утворюється 64 сперматозоїди. Сперматозоїди Фукусмають вічко, розташоване всередині редукованого хлоропласту. У передньому кінці клітини сперматозоїда 13 мікротрубочок формують хобострум- особливу структуру, яка, як припускають, бере участь у визначенні жіночого статевого атрактанта.
Сперматозоїди залучаються до яйцеклітини Фукуснеспецифічним феромоном фукосерратеном, що виділяється яйцеклітиною. Впізнавання ж на видовому рівні пов'язане зі специфічними олігосахаридами на яйцеклітині та сперматозоїді. На поверхні яйцеклітини є різні ділянки, кожна з яких містить особливі органічні сполуки - глікопротеїни. Сперматозоїди містять глікопротеїни, організовані в ділянки на мембрані та виростах переднього джгутика, а також на поверхні сперматозоїда. При зустрічі з яйцеклітиною сперматозоїд рухається над оболонкою яйцеклітини, ніби досліджуючи її за допомогою свого переднього джгутика. Якщо глікопротеїни сперматозоїда комплементарні глікопротеїнам яйцеклітини, тільки тоді відбувається впізнавання сперматозоїду яйцеклітиною.
Систематика
Викопні знахідки, які можуть стосуватися бурих водоростей, відомі із середнього силуру (близько 425 млн. років). Але ці знахідки мають схожість і з деякими сучасними зеленими та червоними водоростями. Викопні знахідки, які можна пов'язати з сучасними бурими водоростями, відносяться до міоцену (5–25 млн. років). Ці водорості нагадують сучасну Диктіоту. Вік бурих водоростей, який визначається молекулярними методами, становить 155–200 млн. років.
Тип організації талому, наявність або відсутність піреноїду, спосіб росту, тип статевого розмноження (ізогамія, гетерогамія, оогамія) та життєвого циклу використовують для виділення порядків бурих водоростей. Останніми роками активно переглядається система бурих водоростей. У різних класифікаційних схемах у складі класу бурі водорості виділяють від 7 до 14 порядків.
Порядок Десмарестієві–Desmarestiales. Водорості кущисті, багаторазово розгалужені, з валькуватою, плоскою або сплощеною слані. Прикріплюється до субстрату добре розвиненою підошвою. На поверхні слані розвиваються волоски.
Рід Десмарестія (рис.33). Бурі або оливкові кущі з добре вираженою головною втечею, до 0.8-1 м висоти. Прикріплюється конусоподібною підошвою. Розгалуження 3–4 порядків, почергове чи супротивне. Гілки останніх порядків плоскі або, рідше, ниткоподібні. Види морські, мешкають в арктичних та помірних широтах на глибині 1-20 м, утворюють зарості або ростуть одинично.

Мал. 33. Десмарестія (Р. Desmarestia)
Порядок Ральфсієві–Ralfsiales. Представники порядку мають слоевище у вигляді кірок різної будови або складаються з базальної кірки та вертикальних пагонів.
Рід Аналіпус.Листок складається з багаторічної кіркоподібної базальної частини і однорічних, оливково-бурих, вертикальних пагонів, що відходять від неї, висотою до 40 см. Багатогніздні спорангії циліндричні за формою і дворядні за будовою. Одногніздні спорангії овальні, розвиваються серед асиміляційних гілок. Види роду широко поширені у морях помірних широт, утворюють пояси у припливно-відливній зоні.
Порядок Диктіотові–Dictyotales.Для видів порядку характерний апікальне зростання. У вегетативних клітинах є численні дископодібні хлоропласти без піреноїдів. Таллом паренхіматозний, характерна ізоморфна зміна поколінь. Статевий процес оогамний, безстатеве розмноження нерухомими спорами, сперматозоїди мають лише один джгутик. Джгутик довгий, з перистими виростами і спрямований уперед. Представники порядку зустрічаються переважно у тропічних і, рідше, в помірних морях.
Рід Диктіота (додаток, 4А). Має слоєвище у вигляді плоских оливково-бурих дихотомічно розгалужених кущиків заввишки до 11-15 см. Гілки лінійні, вузькі, не мають жилок, з роздвоєними вершинками. Тетраспорангії утворюють на поверхні слані соруси, які мають невизначену або кільцеподібну форму. Поширений у морських водах помірних та теплих широт.
Порядок Ламінарієві–Laminariales. Ламінарієві мають слоевище, що складається з пластини, стволика та органів прикріплення – підошви або різозідів. Пластина рівна або зморшкувата, цільна або розсічена, з отворами, складками, булями або без них, завдовжки від кількох десятків сантиметрів до 20-40 м. Стволик розгалужений або частіше нерозгалужений. Спорофити однорічні або багаторічні, вік деяких видів налічує 11-18 років. У корі багатьох ламінарієвих розташовані слизові оболонки.
Найбільш популярними представниками бурих водоростей є ламінарії (види пологів Ламінарія або Сахаріна), . Продукти фотосинтезу – специфічний вуглевод
Рід Сахаріна (Ламінарія).Листок довжиною від 0,5 до 5-10 м, має товсту блискучу пластину, пружний стволик і густо переплетені різоїди, якими прикріплюється до різних твердих субстратів. Краї пластини гладкі або хвилясті, на поверхні пластини часто є опуклості та складки. Скупчення спорангіїв – соруси – утворюються по обидва боки пластини, після виходу суперечка верхня частина пластини руйнується. Живе від двох до кількох років. Сахаріни є цінною промисловою сировиною та об'єктом марікультури. Сахаріна японськаутворює зарості у північній половині Японського моря, а також біля південного та південно-східного берега Сахаліну та біля Південних Курильських островів. Сахаріна японськає найбільш цінним промисловим видом, вона відрізняється чудовими смаковими якостями та використовується для приготування різноманітних страв, консервів, кондитерських виробів.
Порядок Сфацелярієві–Sphacelariales.
Бурі водорості, що відносяться до цього порядку, мають таллом, що гілкується, з апікальним зростанням. Гілки з волосками. Вегетативні клітини мають численні хлоропласти без піреноїдів. Життєвий цикл у більшості з ізоморфною зміною поколінь, статевий процес – ізогамія або гетерогамія. Водорості утворюють невеликі бурі пучки до 3 см у висоту та розгалужені кущики до 20 см у висоту на камінні або інших водоростях; є ендофітні представники.
Рід Сфацелярія. Має тонконітчасте слоевище. Нитки, що гілкуються, утворюють невеликі, до 0,5-3 см висоти, пучочки бурого кольору. Вегетативне розмноження здійснюється спеціалізованими, часто трипроменевими вивідковими гілочками. пропагулами. Статевий процес ізогамний. Жіночі гамети виділяють феромони ектокарпен та мультифіден. Види роду поширені у морях як полярних, і тропічних широт.
Порядок Сцитосифонові–Scytosiphonales. Представники з кишкоподібною, міхуровою, пластинчастою або мішковидною, часто порожнистою слані до 25 см довжини. Кора однорядна або дворядна, внутрішня частина (стінка) слані утворена кількома шарами великих клітин. На поверхні слані розвиваються багатогніздні спорангії, які зібрані в соруси, що підтримують їх нитки. парафізита пучки волосків.
Рід Сцитосифонє тонкостінними бурими трубками 4–25 см довжини (додаток, 4Б). Листок прикріплюється невеликою підошвою і має рідкісні поперечні перетяжки. Зустрічається у прибережній зоні морів у різних широтах, часто утворює пояси у припливно-відливній зоні.
Порядок Фукусові–Fucales. Мають багаторічну слоевище у вигляді великих кущів, що неправильно або дихотомічно гілкуються, до 1-3 м довжини. Прикріплюються до субстрату конусоподібною підошвою. Слані мають пагони, філоїди (листкоподібні утворення), рецептакули (містилища гамет) і плавальні (повітряні) бульбашки. Зустрічаються в припливно-відливній зоні всіх помірних та арктичних морів, де утворюють потужні чагарники – пояси.
Рід Саргассум(Рис. 34, У). Рясно розгалужені оливково-бурі або коричневі кущі заввишки 05-15 м. Прикріплюються конічною підошвою. Філоїди розвиваються на бічних пагонах, що відходять від циліндричного стовбура, в нижній та середній частинах слані. Філоїди шкірясті, з центральним ребром, 0,2-2 см довжини. Плавальні пухирі овальні або округлі, з виростами або без них. Саргасси широко поширені в помірних та субтропічних широтах від нижньої літоралі до глибини 10 м, де утворюють щільні або розріджені чагарники. У південній частині Тихого океану – так званому Саргасовому морі – утворюють великі плаваючі скупчення, у яких розмножуються лише вегетативним шляхом.
Рід Фукус (рис.32).Словище шкірясте, кущисте, розгалужується дихотомічно, висотою 25-50 см. Біля узбережжя далекосхідних морів росте кілька років, мешкає на літоралі. Часто утворює великі зарості на мілководді. Для видів цього роду характерно дихотомічно

Мал. 34. Зовнішній вигляд фукусових водоростей: А – Аскофілум; Б – Цистозіра; У – Саргассум; 1 – рецептакул, 2 – плавальний міхур
розгалужена слоевище з плоскими гілками, що мають поздовжнє ребро, яке прикріплюється до каменів конічної підошвою. Фукуси використовують як добрива, як корм для худоби, для виробництва кормового борошна, альгінатів, застосовують у медицині для лікування зоба, корекції ваги і т.д.
Порядок Ектокарпові–Ectocarpales. Водорості, які сюди належать, мікроскопічні, вільноживучі, епіфіти, епізоїти чи ендофіти. Органи розмноження представлені одногніздними (спорангії) та багатогніздними (гаметангії) спорангіями; вони довгоциліндричні, короткоциліндричні або стручкоподібні.
Рід Ектокарпус. Лист у вигляді м'яких бурих кущиків до 2 см висоти. Кущики утворені однорядними, по черзі, дихотомічно або неправильно розгалуженими нитками. Багатогніздні спорангії стручкоподібні, утворюються збоку бічних гілок на 1-3-клітинній ніжці. Види поширені в морях різних широт, у чистих та забруднених водах, на літоралі та у верхній частині субліторалі. Епіфіти зустрічаються і в обростанні антропогенних субстратів.
Екологія
Переважна більшість представників бурих водоростей живе у морських водоймах. У прісних водах трапляються лише 8 видів. Трапляються як однорічні, так і багаторічні види, вік яких може досягати 15-18 років. Ламінарієві водорості Нереоцистісі Макроцистис, Які мешкають вздовж тихоокеанського узбережжя Північної Америки, мають максимальні для всіх водоростей розміри - до 30-40 м завдовжки. Вони формують гігантські підводні ліси морях Далекого Сходу. Прикріплюються бурі водорості до найрізноманітніших субстратів – скель, каменів, гравію, раковин та панцирів морських тварин, інших водоростей. Деякі дрібні форми бурих водоростей живуть усередині тканин інших водоростей як ендофіти. Більшість видів бурих живуть у прикріпленому стані. Відірвані від ґрунту слані зносяться течією у спокійні місця з мулистим дном і продовжують там існувати. Види з повітряними бульбашками на слоевище, відірвавшись від ґрунту, спливають до поверхні, утворюючи великі плаваючі скупчення, особливо в районах зі стабільною круговою течією, як у Саргасовому морі. Вони існують у таких скупченнях досить довгий час і розмножуються лише вегетативним способом.
Бурі водорості широко поширені у всіх морях, але найбільшого розвитку вони досягають у морях помірних та приполярних широт. Їхні великі слані ростуть, прикріпившись до скель і каменів, а в спокійних місцях біля берега і на великій глибині можуть утримуватися навіть на стулках раковин молюсків та гравії. Зустріти їх можна на різній глибині – від літоральної зони, де під час відливу вони годинами знаходяться поза водою до 40–200 м. У районі Гавайських островів на глибині 180 м виявлено види роду Саргассум, а Ламінаріяв Адріатичному морі було знайдено на глибині 200 м. Найбільш рясні зарості бурих водоростей спостерігаються на глибині 6–15 м, де найкращі умови освітлення та постійний рух води, що приносить до їх слоїв біогенні речовини та обмежує поселення рослиноїдних тварин. фітофагів.
Золотисті водорості –Chrysophyceae
Одноклітинні, колоніальні або багатоклітинні організми, які зазвичай мають джгутикову стадію в життєвому циклі. Клітини одноядерні, з одним або декількома хлоропластами. Стигма зазвичай є (рис. 36). Жгутиків 1-2, нерівних. Скорочувальна вакуоль розташована в передній частині клітини. Зовнішні покриви клітин цих організмів дуже різноманітні. У найбільш примітивних видів клітина одягнена ніжним перипластом, що дозволяє утворювати випинання різної форми (ризоподії, псевдоподії), в інших плазмалемму покриває тверда целюлозна оболонка, що іноді сильно ослизняється. Будиночки у золотистих водоростей бувають різної форми: вазоподібні, кулясті, яйцеподібні, циліндричні; з одним або декількома отворами. Клітина прикріплюється до основи будиночка за допомогою гнучкої ніжки або може бути вільною. У деяких хризофіт на поверхні клітини відкладаються вапняні утворення - коколіти - різних форм і розмірів, а кремнієві джгутиконосці мають внутрішній кремнієвий скелет. Різноманітність будови цих скелетів дозволяє використовувати залишки тих чи інших форм викопних силікофлагелят для визначення віку геологічних порід, що їх містять. У більш високорозвинених представників відділу золотистих водоростей клітини одягнені панциром, що складається з кремнієвих лусочок (іноді несучі шипи), або укладені в будиночки, через отвори яких назовні виходять джгутики або псевдоподії.
– поперечний зріз стволика;
Пігменти – хлорофіли аі з, фукоксантін. Для хризофіт характерний найвищий серед гетероконтів вміст фукоксантину (55-93% від усіх каротиноїдів).
Більшість видів - прісноводні фототрофи, але зустрічаються гетеротрофи та фаготрофи.
Запасні речовини – хризоламінарин та олії.
Серед одноклітинних форм золотистих водоростей переважають прісноводні планктонні види, серед багатоклітинних – донні чи эпифитные. Зустрічаються в основному в прохолодних морських та прісних водоймах. Відомо близько 360 видів.

Мал. 36. Золотисті водорості (по: С. Hoek van den et al., 1995): А– будова клітини; Б- Корінецька система; У- Тричастинна мастигонема; Г- Зріз частини хлоропласту; 1 – базальне тіло; 2 – базальне здуття; 3 – короткий джгутик; 4 – стигма; 5 – ризопласт; 6 – ядро; 7 – ядерна мембрана; 8 - ядерце; 9 – мітохондрія; 10 – вакуолі; 11 - слиз; 12 – плазмалема; 13 - везикули; 14 – слизове тіло; 15 – ліпіди; 16 – хлоропласт; 17 - апарат Гольджі; 18 – скорочувальна вакуоля; 19 - довгий джгутик: 20 - 1-й корінець; 21 - 2-й корінець; 22 - 3-й корінець; 23 - 4-й корінець; 24 - хлоропластний нуклеоїд; 25 - ХЕС; 26 – оболонка хлоропласту; 27 - оперізує ламелла; 28 - ламелла; 29 – довгий латеральний філамент; 30 – короткий латеральний філамент; 31 – термінальний філамент; 32 - трубчаста частина мастигонеми
Структура таломуу золотистих водоростей найчастіше монадна, але може бути дуже різноманітною: кокоїдною, пальмелоїдною, плазмоїдальною, амебоїдною, нитчастою, паренхіматозною (рис. 37).

Мал. 37. Золотисті водорості (по: Л.Л. Великанов та ін, 1981): А – Охромонас: 1 – зовнішній вигляд; 2 – циста; Б – Хромуліна: 1 – зовнішній вигляд; 2 – циста; 3 – схема утворення плівки з цист; У – Дінобріон: 1 – загальний вигляд колонії; 2 – циста; Г – Хризамеба; Д – Гіббердія; Е – Гідрурус: 1 – загальний вигляд колонії, 2 – кінчик гілки, 3 – зооспора
Розмноження. У золотистих водоростей зустрічаються три способи розмноження: вегетативне, безстатеве і статеве.
Вегетативне розмноженнявідбувається шляхом поздовжнього поділу клітини або розпаду колонії на частини за рахунок поділу клітин в одному, двох або трьох напрямках.
Статевий процес(клітини золотистих водоростей диплоїдні) - ізогамія або кон'югація. В результаті статевого процесу, а також у несприятливих умовах утворюються цисти з товстою оболонкою, що містить кремній.
Безстатеве розмноженняздійснюється одно- або двожгутиковими зооспорами, які розвиваються у звичайних вегетативних клітинах або зооспорангіях. Статевий процес: голо-, ізо- та автогамія. Гологамія – вид статевого процесу, при якому дві подібні з вегетативними клітинами зливаються на передніх кінцях у двоядерну зиготу, яка перетворюється потім на окремі цисти.
Систематика
Із Золотистих водоростей наприкінці минулого століття на основі результатів молекулярно-генетичних досліджень виділено клас Синурові. Також виділено клас Феотамнієві – із Жовтозелених та Золотистих водоростей. Тому обсяг класу Chrysophyceae скоротився і в даний час налічує близько 360 видів, що належать до трьох порядків: Chromulinales, Hibberdiales і Hydrurales. Форма, будова та кількість джгутиків, а також пігментний склад та структура тіла є важливими систематичними ознаками.
Порядок Хромулінові – Chromulinales.
Рід ХризамебаДо цього порядку відносять організми з монадним, пальмелоїдним та амебоїдним типами диференціації талому. Монадні клітини з одним джгутиком, видимим у світловий мікроскоп. А(Рис. 37,
Рід ХромулінаДо цього порядку відносять організми з монадним, пальмелоїдним та амебоїдним типами диференціації талому. Монадні клітини з одним джгутиком, видимим у світловий мікроскоп. У) – прісноводні амебоїдні водорості, вони мають у життєвому циклі джгутикову стадію. Зигота перетворюється на стоматоцисту.
Рід Дінобріон) - одноклітинні вільноплаваючі організми, що мешкають переважно в прісних водах. Клітини голі; є одна чи дві золотисті пластиди. (Рис. 37, Г)
– одноклітинні та колоніальні, вільноплаваючі або прикріплені представники. Монади знаходяться у вазоподібних будиночках, утворених целюлозними мікрофібрилами. Крім целюлози до складу будиночка входить і багато амінокислот. Кущикоподібна форма колоній пов'язана зі способом вегетативного поділу клітин, коли одна з дочірніх клітин, залишаючи батьківський будиночок, прикріплюється до його отвору. Обертаючись навколо своєї осі, вона формує власний будиночок. Батьківський будиночок можуть залишити і обидві дочірні клітки. Клітини містять 1-2 хлоропласти, на передньому кінці клітини видно вічко і 2 скорочувальні вакуолі. Хризоламінарин розташований у вакуолі на задньому кінці клітини. Розмноження вегетативне та статеве.–Порядок Гіббердієві Hibberdiales
. Для представників цього порядку характерний унікальний пігментний склад. Крім фукоксантину, вони мають додатковий світлозбираючий каротиноїдний пігмент - антераксантин. Включає пологиГіббердія, Хромофітон, Стилоцерас, Хризопіксис, Платитека
Рід Гіббердіята інші.
має дві стадії в життєвому циклі: колоніальну пальмелоїдну нерухому і одноклітинну рухливу монода. При світловій мікроскопії у монадної стадії видно лише один джгутик. Характерна наявність крім фукоксантину ще одного додаткового світлозбираючого каротиноїдного пігменту – антераксантину.–Порядок Гідрурусові. Представники даного порядку мають таллом з пальмелоїдним та псевдопаренхіматозним типами будови. Характерна наявність унікальних зооспор тетраедричної форми. Ока немає. Мітоз напівзакритий.
. Для представників цього порядку характерний унікальний пігментний склад. Крім фукоксантину, вони мають додатковий світлозбираючий каротиноїдний пігмент - антераксантин. Гідрурус, Целлонієлла, Федерматіум, Хрізонебулата інші.
Рід Гідрурус(Рис. 37, Е) є найбільш диференційованим серед пальмелоїдних водоростей. Його талом є великими, довжиною до 30 см слизові колонії, що мають вигляд коричневих розгалужених шнурів, що часто видають дуже неприємний запах. У колоніях можна розрізнити головний стовбур та бічні розгалуження. У клітинах, занурених у загальний слиз, міститься один чашоподібний хроматофор та кілька скорочувальних вакуолей. Клітини рихло розташовані по периферії колонії і щільніше в середині. Таллом може зростати лише за рахунок поділу апікальних клітин. При безстатевому розмноженні у клітинах бічних гілок колонії формуються зооспори. Утворює кулясті цисти. Зустрічається у гірських струмках та річках з холодною водою, де прикріплюється до твердих субстратів.
Клас Синурові водорості –Sinurophyceae
syn- разом і ura– хвіст. Ця група організмів у 1987 р. виділена із золотистих водоростей. Поєднує монадні одиночні та колоніальні організми, іноді з чергуванням у життєвому циклі монадної та пальмелоїдної стадії. Поверхня пектинових оболонок покрита панциром із кремнієвих лусочок. Мітохондрії з трубчастими христами зазвичай розташовані в цитоплазмі навколо хлоропласту. Ядро одне, хлоропластів, оточених чотирма мембранами, зазвичай два. Ламели тритілакоїдні, оперізуюча ламела є. Основні пігменти – хлорофіли аі з, β -каротин та фукоксантин. Стигми немає. Клітини зазвичай з 2 нерівними джгутиками. Довгий перистий джгутик спрямований уперед. Короткий гладкий джгутик, який іноді сильно редукований, спрямований назад.
Клітини розмножуються переважно поздовжнім розподілом. Колонії, розпадаючись, дають початок молодим колоніям. У деяких видів описаний статевий процес у формі ізогамії. При цьому, на відміну від золотистих водоростей, злиття ізогамет у синурових водоростей відбувається не на передніх, а на їх задніх кінцях. Ендогенним шляхом утворюються окремі цисти з часом, подібні з такими у хризофіт.
Переважно планктонні форми, пальмелоїдні стадії входять до складу бентосу. Більшість видів синурових водоростей – фототрофні прісноводні організми.
Систематика.
Клас синурових водоростей включає 7 порядків: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales і Thallochrysidales.
Порядок Синурові – Synurales. Монадні форми, пектинові оболонки клітин зазвичай покриті панциром з кремнеземних лусочок, зцементованих органічною речовиною в цілісний футляр. Хлоропластів один чи два.
Рід Синура(Рис. 38) - прісноводні монадні форми з двома нерівними джгутиками та двома пристінковими хлоропластами. Ядро грушоподібне, розташоване у передній частині клітини. Позаду ядра є одна велика вакуоль, а задній частині клітини – кілька дрібних скорочувальних вакуолей. Клітини покриті лусочками, як черепицею Розмноження найчастіше вегетативне, для деяких відомий статевий процес. Утворює колонії (рис. 39, А). Часто у всіх клітинах колонії виникають цисти. При масовому розвитку Синураможе надавати воді неприємного запаху.

Мал. 38. Зовнішній вигляд клітин Синури
Порядок Ризохлоридові – Rhizochloridales. Переважно прісноводні організми з ризоподіальним типом диференціації талому, що живуть головним чином у прісних водоймах.
Рід Малломонас(Рис. 39, Б-Г) – одноклітинна монада з одним добре помітним джгутиком та одним роздвоєним хлоропластом.

Мал. 39. Зовнішній вигляд синурових водоростей (по: Г.А. Белякова та ін., 2006): А – колонія, що ділиться Синури; Б, В – освіта статоспори та Г – зовнішній вигляд Малломонаса
Клітина покрита лусочками черепітчасто і по спіралі, деякі з них несуть голкоподібні шипи (рис. 39, Г). Характерна наявність статоспор (рис. 39, Б, В). Для низки представників описаний статевий процес – гологамія.
Рід Міксохлорісмешкає в клітинах листя сфагнових мохів, має вигляд великого багатоядерного плазмодія. Восени він утворює цисти, які проростають навесні. З них виходять зооспори або амеби, що проникають у порожні клітини листя і зливаються там у плазмодії.
Порядок хлоромебових - Chloramoebales.Включає монадних представників. Зустрічаються у солоних та прісних водах.
Рід Гетерохлоріс – його клітини можуть змінювати форму, утворюючи псевдоподію. Це явище притаманне багатьом представникам порядку, як і тенденція до тваринного способу харчування. У клітці є кілька хлоропластів, краплі олії та хризоламінарин. У передній частині клітини є скорочувальні вакуолі, у центрі – одне ядро. Розмножується вегетативно поздовжнім поділом клітини.
Порядок Охромонадові - Ochromonadales. Поєднує голі форми з двома нерівними джгутиками. Прісноводні та морські форми.
Рід Охромонасвключає одноклітинні монади з двома нерівними джгутиками. Клітини одягнені лише плазмалемою. На передньому кінці клітини є скорочувальна вакуоля та вічко, на задньому кінці – вакуоля з хризоламінарином. Розмножується вегетативно, борозна розподілу починається біля переднього кінця клітини між двома парами джгутикових основ. Види роду поширені в оліготрофних прісних водах, але є і морські представники.
Порядок Гетероглеєві - Heterogloeales. Включає водорості з пальмелоїдним типом диференціації талому. Представники порядку зустрічаються частіше у прісних, ніж у солоних водах.
Рід Гельмінтоглеямешкає в солонуватих водах. Ця колонія, що сидить на розширеній підставі, складається з слизових тяжів, що гілкуються. У цьому слизу безладно розташовуються протопласти, кожен із яких оточений власної слизової обгорткою.
Клас Феотамнієві водорості – Phaeothamniophyceae
Назва класу походить від типового роду Phaeothamnion(Від грец. phaeos- Смаглявий кущик). Представників цієї групи організмів виділили в окремий клас із жовтозелених та золотистих водоростей у 1998 р. на підставі аналізу послідовності рибосомальних генів та особливостей ультраструктури клітини. Для феотамнієвих водоростей характерна унікальна комбінація пігментів: фукоксантину з гетероксантином, відсутність віолаксантину.
Особливості будови клітини: відсутність вакуолей із хризоламінарином та відсутність ендогенних цист із кремнеземними стінками.
Клас включає одноклітинні, колоніальні та багатоклітинні організми з кокоідним, пальмелоїдним та нитчастим типом структури талому. Жгутикові стадії з двома нерівними джгутиками. Джгутики латеральні або субапікальні. Стигма зустрічається у зооспорі. Пігменти – хлорофіли аі з, β -каротин, фукоксантин, діадіноксантін, діатоксантин та гетероксантин. Основним запасним продуктом є парамілон ( β -Глюкан). Клітинна стінка є, під час клітинного поділу дочірня стінка формується усередині батьківської стінки.
У феотамнієвих з основних способів розмноження зустрічається лише вегетативне та безстатеве. Безстатеве розмноження здійснюється автоспорами або зооспорами. Статеве розмноження невідоме.
Види Феотамнієвих водоростей мешкають виключно у прісних водах. Систематика.
В даний час підтверджена монофілетичність класу феотамнієвих водоростей, і серед охрофіту він ближчий бурим і жовтозеленим водоростям, ніж золотистим. В даний час відомо близько 30 видів цих водоростей, що належать до одного порядку Феотамнієві – Phaeothamniales.
Рід Феотамніонявляє собою прикріплені нитки, що гілкуються, до 1 см висоти (рис. 40). Клітини циліндричні, неправильні, розширені догори,

Мал. 40. Зовнішній вигляд Феотамніону.
довжині вдвічі-втричі перевищують ширину. При розподілі клітин матеріал батьківської стінки витрачається на утворення шаруватого чохла, що оточує клітини нитки. По периферії клітини під плазмалемою є везикули. Вони нагадують фізиди – утворення, що зустрічаються у клітинах бурих водоростей. Клітини містять від однієї до кількох дископодібних пластид оливково-бурого кольору. Зооспори утворюються по 1-2, рідше по 4-8 в одній клітині. У зооспор джгутики прикріплюються збоку. Положення джгутикових корінців у Феотамніонунагадує таке у жовтозелених та бурих водоростей. У нього також відсутня ризопласт. У життєвому циклі можуть утворюватися цисти, але у стінках відсутня кремній. Цисти проростають зооспорами. Поселяється як епіфіт на ниткових водоростях. Мешкає у стоячих та повільно поточних прісних водоймах.
Клас Рафідові водорості –Raphidophyceae
Назва класу походить від грец. rhaphid - Голка. Поєднує одноклітинні двожгутикові організми, позбавлені клітинної стінки. В основному прісноводні організми із сплощеними клітинами (рис. 41).
Клітини мають джгутики нерівної довжини, спрямований вперед джгутик - перистий і довше, спрямований назад - гладкий і коротший. Перехідна спіраль відсутня. Хлоропласти дрібні, численні чи, рідше, по 1-2 у клітині, оточені чотирма мембранами, їх дві мембрани ХЕС. Тилакоїди зібрані в стоси по три. Ядро велике, оточене кільцем диктіосом. Мітохондрії з трубчастими христами. Є скорочувальні вакуолі. Цитоплазма часто буває вакуолізованою. Клітини голі, оточені лише плазмалемою. Стигми (вічка) зазвичай немає.
Фотосинтетичні пігменти – хлорофілі аі з, β -каротин, вошеріаксантін, діноксантін, діадіноксантін, гетероксантин.
Запасні продукти асиміляції – жири та олії, рідше крохмаль та хризоламінарин.
Мешкають також у сфагнових болотах та інших місцеперебуваннях з кислою та нейтральною реакцією води. Деякі види зустрічаються в солонуватих та морських водах.

Мал. 41. Зовнішній вигляд рафідової водорості.
Систематика.
Дуже довго цю групу водоростей через зелене забарвлення, наявність глотки та ряд інших особливостей будови розглядали як клас Хлоромонади серед Євгленових водоростей. Але дані цитології, біохімії, фізіології та молекулярної біології показали їхню безперечну приналежність до відділу Гетероконтні (Охрофітові) водорості та монофілетичність групи Рафідових водоростей.
Відомо близько 25 видів з єдиного порядку Хаттонелові– Chattonellales.
Рід Гетеросигма(Рис. 42, А) включає морські флагеллати. Клітини овальної форми зі джгутиками, зрушеними на бік, пластиди розташовані по периферії клітини. Трихоцист немає. Плавають, здійснюючи обертові рухи навколо поздовжньої осі тіла. Можуть викликати токсичне "цвітіння води" у прибережних водах.

Мал. 42. Рафідові водорості (по: R. E. Lee, 1999): А – Гетеросигма; Б – Гоніостомум; 1 – хлоропласт; 2 – мукоциста; 3 – скорочувальна вакуоля; 4 – трихоциста; 5 – ядро
Рід Гоніостомум(Рис. 42, Б) - рухливі дорсовентрально стислі монади. Спинна сторона вигнута, черевна – сплощена. Борозна на передньому кінці веде в трикутну горлянку, з якої виходять два довгі джгутики; їх довжина можна порівняти з довжиною клітини. Пластиди розташовані на периферії клітини. Під плазмалемою знаходяться трихоцисти. Мешкає в прісних водах із кислотним значенням рН.
Клас Евстигмові водорості –Eustigmatophyceae
Назва класу походить від грец. eu- "добре та stigma-«Мітка», «пляма». Клас поєднує голі одноклітинні, рідше колоніальні, організми переважно кокоідної структури.
Ядро одне чи кілька. Хлоропласт зазвичай один, оточений 4 мембранами. Джгутик зазвичай один передній перистий і друге базальне тіло або, рідше, два нерівні джгутики. Стигма (вічко) є, розташована за межами хлоропласту, її гранули без мембран.

Мал. 43. Зовнішній вигляд евстигмової водорості
Хлоропласт зазвичай єдиний, великий, багатолопатевий, чашоподібний або парієтальний, жовто-зеленого кольору. Тилакоїди зібрані в ламелли по три, оперізувальна ламелла відсутня.
Основні пігменти: хлорофіла, β-каротин, віолаксантин і вошеріаксантин. Піреноїд зазвичай є тільки у вегетативних форм.
Запасні речовини – олії та речовина невстановленої природи, яка відкладається у вигляді твердого матеріалу поза пластидами. Крохмаль не утворюється.
Мешкають у прісних водах, рідше зустрічаються в морях та у ґрунті.
Систематика.
Раніше цю групу організмів відносили до Жовтозелених водоростей. Нині Євстигмові водорості розглядають у ранзі класу у відділі Гетероконтофіти. Відомо близько 35 видів, переважно прісноводних, що належать до єдиного порядку Євстигмові – Eustigmatales.
Рід Євстигматос(Рис. 43) - одноклітинні дрібні кулясті водорості. Клітинна стінка гладка, цілісна, без орнаменту. Жовто-зелений хлоропласт один, лопатевий, парієтальний, зі стебловими багатогранними піреноїдами. Велика центральна вакуоля з червоним вмістом. Розмножується 2 або 4 автоспорами або бутілеподібними зооспорами з одним вхідним джгутиком, великою стигмою, розташованою за межами хлоропласту на передньому кінці, і єдиним заднім хлоропластом без піреноїду. Звичайний компонент ґрунтових флор. Відомий з Нової Зеландії, Австрії, Ісландії, та з басейнів Арізони як "гірчична водорість".
Клас Жовтозелені водорості –Xantophyceae
До Жовтозелених відносяться водорості, хлоропласти яких забарвлені у світло-або темно-жовтий колір, дуже рідко зелений і лише іноді блакитний. Забарвлення таломів визначається наявністю в хлоропластах клітин наступних пігментів – хлорофілів аі з, β -каротину та ксантофілів. Переважна більшість останніх і зумовлює своєрідність забарвлення жовтозелених водоростей. Крім того, в клітинах як основний продукт асиміляції накопичуються парамілон, краплі олії і лише у деяких видів, крім того, глибки лейкозину і волютину. Крохмаль у жовтозелених водоростей не утворюється. Відмінною особливістю жовтозелених є наявність у вегетативних клітин монадної структури та у зооспор двох нерівних джгутиків. До складу клітинної стінки входять целюлоза, глюкоза та уронові кислоти. Клітинна стінка часто складається із двох частин.
Розмноження вегетативне, безстатеве та статеве.
Широко поширені у прісних водах. Рідко зустрічаються в морських, солонуватих водах та у ґрунті.
Раніше клас Жовтозелених водоростей називався Трібофіцієві водорості за типовим родом Трібонема (від грец. tribon – майстерний, хитрий і nema – нитку). Відомо близько 450 видів.
Жовтозелені характеризуються значною морфологічною різноманітністю. Серед численних представників цього відділу виявляються майже всі основні типи структури тіла: амебоїдна, монадна, пальмелоїдна, кокоїдна, нитчаста, різнонітчаста, пластинчаста та сифональна (рис. 44). – 46). Словник одноклітинний,

Мал. 44. Зовнішній вигляд жовто-зелених водоростей: 1, 2 – Хараціопсис, 3 – Центритрактус, 4 – Офіоцитіум
колоніальне, багатоклітинне та неклітинне. Клітинна оболонка щільна, пектинова і целюлозна, що складається з частин, що щільно налягають один на одного, або з двох стулок. В оболонці відкладається кремнезем чи вапно. Переважно нерухомі форми. Серед одноклітинних видів є рухомі форми, позбавлені щільної оболонки та забезпечені джгутиками, лобоподіями та різоподіями.

Мал. 45. Зовнішній вигляд ксантококкових жовто-зелених водоростей: 1–3 – Ботрідіопсис, 4 – Тетраедрієлла, 5 – Псевдостаураструм, 6 – Гоніохлоріс, 7, 8 – Бумілеріопсис
Більшість жовтозелених – нерухомі організми. У рухомих особин рух може здійснюватися за допомогою джгутиків чи різоподій. Клітини різної форми: кулясті, веретеноподібні, еліпсоїдні, циліндричні, тетраедричні, серпоподібні, грушоподібні, яйцеподібні. Талломи розмірами від 0.5 – 1.5 мкм ( Хлорідела) до кількох міліметрів у діаметрі ( Ботрідіопсис) (рис. 45, 1 – 3) і до десятків сантиметрів завдовжки ( Вошерія) (рис. 46, 3).

Мал. 46. Зовнішній вигляд жовто-зелених водоростей: 1 – Трибонема, 2 – Гетеропедія, 3 – Вошерія, частина нитки з оогонієм та антеридієм
Більшість видів жовтозелених є фототрофами, але трапляється і голозойне харчування шляхом заковтування бактерій та дрібних водоростей. Жовтозелені водорості широко поширені у прісних водах. Вони також звичайні в ґрунті, рідше зустрічаються в морських та солонуватоводних водоймах. Клас включає аеробіонтні, планктонні, донні та перифітонні форми. Епіфіти, епізоїти, а також внутрішньоклітинні симбіонти у найпростіших клітинах.
Незалежно від зовнішньої структури, внутрішня будова клітини жовто-зелених водоростей однотипна. У протопласті спостерігається зазвичай кілька жовто-зелених хлоропластів, що мають дископодібну, коритоподібну, пластинчасту, рідше стрічкоподібну, зірчасту або чашоподібну форму з цільними або лопатевими краями. Забарвлення пов'язане з відсутністю фукоксантину, що відповідає за золотистий та коричневий колір в інших охрофітових. З інших пігментів вони мають β -каротин, вошеріаксантин, діатоксантин, діадіноксантін, гетероксантин. У рухомих форм на передньому кінці хлоропласту зазвичай розташоване червоне вічко, або стигма. У небагатьох видів є піреноїди напівзавантаженого типу. Ядро в клітині зазвичай одне, невеликих розмірів, але є види і з багатоядерними клітинами. У деяких видів у передній частині клітини є одна або дві скорочувальні (пульсуючі) вакуолі.
У монадних представників і рухомих стадій (зооспор і гамет) є два нерівних джгутика. Винятком є синзооспори Вошерії, у яких по поверхні розташовані численні пари трохи розрізняються по довжині гладких джгутиків. Короткий джгутик закінчується акронемою. Джгутики прикріплюються на клітині субапікально. У сперматозоїда Вошеріїприкріплення латеральне.
У видів з амебоїдною, монадною та пальмелоїдною організацією клітинна стінка відсутня, вони покриті лише цитоплазматичною мембраною і можуть легко змінювати форму. Іноді «голі» клітини знаходяться усередині будиночків, стінки яких можуть бути забарвлені у бурий колір солями марганцю та заліза. У більшості форм є клітинна стінка, що складається з двох частин. У складі клітинної стінки переважає целюлоза, а також містяться полісахариди, що складаються в основному з глюкози та уронових кислот. У молодих клітин оболонка тонка, з віком потовщується. У ній можуть відкладатися солі заліза, з'єднання якого забарвлюють її у різні відтінки коричневого та червоного тонів. Найчастіше в клітинній стінці є кремнезем, надаючи їй твердість і блиск. Вона може інкрустуватися також вапном і бути по-різному скульптурована (шипики, осередки, бородавки, щетинки, зубчики і т.д.) У прикріплених форм може утворюватися виріст оболонки – ніжка з прикріпною підошвою.
У нитчастих форм жовтозелених водоростей із двостулковими оболонками при розпаді ниток клітинні оболонки розвалюються на Н-подібні фрагменти. Ці фрагменти є щільно з'єднаними половинками оболонок двох сусідніх клітин (рис. 47). При зростанні ниток Н-подібний фрагмент клітинної стінки двох сусідніх дочірніх клітин вбудовується між двома половинками материнської оболонки клітини. У результаті кожна з дочірніх клітин наполовину покрита старою оболонкою материнської клітини та наполовину. – новоствореною оболонкою.

Мал. 47. Схема утворення поперечної перегородки між двома дочірніми клітинами у нитчастих жовто-зелених водоростей (по: А.А. Масюк, 1993): А- Фрагмент нитки; Б- Закладка пояскового кільця та утворення поперечної перегородки між двома клітинами; У– шаруватість двостулкових клітинних оболонок; Г- Розпад оболонки на Н-подібні ділянки
Скорочувальні вакуолі є у рухливих представників. Зазвичай їх 1-2 на клітину. Апарат Гольджі своєрідної будови. Диктіосоми дрібні, містять 3-7 цистерн. Ядро одне, рідше їх багато; у ценотичних видів клітини завжди багатоядерні.
Розмноження. Для більшості видів жовтозелених водоростей характерно вегетативне та безстатеве розмноження.
Вегетативне розмноженняздійснюється різними способами: розподілом клітин навпіл, розпадом колоній та багатоклітинних слоевищ на частини. У Вошеріїутворюються спеціальні виводкові нирки.
При безстатевому розмноженніможуть формуватися різноманітні суперечки: амебоїди, зооспори, синзооспори, автоспори, гемізооспори, геміавтоспори, апланоспори. Зооспори «голі» і зазвичай грушоподібної форми.
Статевий процес– ізогамія, гетерогамія та оогамія – описаний у небагатьох представників. У Трибонемигамети подібні за розмірами, але різняться поведінкою – це ізогамія. У Вошеріїспостерігається оогамія: на нитках утворюються вмістилища жіночих гамет – оогонії та чоловічих – антеридії.
У несприятливих умовах спостерігається утворення цистів. Цисти (статоспори) ендогенні, одноядерні, рідше багатоядерні. Їхня стінка часто містить кремнезем і складається з двох нерівних або, рідше, рівних частин.
Систематика.
Наприкінці XIX – на початку XX ст. різні пологи жовтозелених відносили до відділу зелених водоростей, що було пов'язано в першу чергу з забарвленням та морфологічною схожістю таломів. В даний час жовтозелені розглядаються як клас усередині відділу охрофітових.
Відомо близько 450 сучасних видів класу Жовтозелених водоростей, які групуються в чотири порядки: Ботрідієві, Мішококкові, Трибонемові та Вошерієві. В основу виділення порядків покладено тип диференціації талому та особливості життєвого циклу.
Порядок Ботридієві – Botrydiales. До порядку відносяться види з сифональним типом диференціації талому, у яких відсутній оогамний статевий процес.
Рід Ботрідіуммешкає на ґрунті і має вигляд зелених пухирів завбільшки кілька міліметрів, прикріплених за допомогою безбарвних ризоїдів. Таллом сифональний, містить численні ядра та пластиди. Оболонка багатошарова, на ній може відкладатися вапно. Розмноження безстатеве за допомогою двожгутикових зооспор, при цьому весь вміст міхура розпадається на одноядерні фрагменти. При нестачі вологи розмножується за допомогою апланоспор або формує товстостінні цисти. В одних випадках весь вміст міхура йде на утворення однієї великої цисти. В інших випадках цисти формуються в ризоидах, куди спочатку переходить вміст міхура. Цисти проростають або у новий таллом, або утворюють зооспори. Статевий процес - ізо-і гетерогамія. Зигота проростає негайно, без періоду спокою. Звичайні та широко поширені в наземних місцеперебуваннях види, що зустрічаються по берегах струмків, ставків або на позбавлених рослинності ґрунтах.
Порядок Місхококові–Mishococcales. Одноклітинні або колоніальні представники з кокоїдним типом диференціації талому.
Рід Хараціопсисвключає прикріплені одноклітинні форми. При розмноженні формує зооспори, апланоспори та товстостінні цисти (рис. 44, 1-2).
Рід Офіоцитіум(Рис. 44, 4) має подовжені циліндричні клітини, які можуть бути прямими, зігнутими або спірально закрученими, а на кінці можуть нести шип. Клітинна стінка і двох нерівних частин, їх більшість бере участь у зростанні клітини, менша частина стала і має форму кришечки. Одноклітинні та колоніальні види, що вільно живуть або прикріплені до субстрату за допомогою маленької ніжки. Розмножуються зооспорами та апланоспорами, зустрічаються цисти. Мешкають у прісних водах.
Рід Місхококкусутворює деревоподібні прикріплені колонії. Розгалуження дихотомічне та тетрахотомічне. Клітини розташовані по 2 чи 4 на верхівках слизових гілок колонії. Клітини кулясті, до овальних, з тонкою або товстою клітинною стінкою. Іноді клітинна стінка блискуча та коричнева через просочення її солями заліза. Молоді одноклітинні організми зі слизовою дископодібною основою, яка служить точкою опори при прикріпленні. Після виходу суперечка протопласт материнської клітини перетворюється на холодець і витягується, довжина стає в 6 разів більшою за ширину, і таким чином виникає циліндрична ніжка. Порожня клітинна стінка материнської клітини завжди стає основою ніжки. Безстатеве розмноження зооспорами та автоспорами. Автоспори прикріплюються до верхнього краю слизової оболонки ніжки. Наступні клітинні поділу повторюють процес і виробляють деревоподібну колонію. Статевий процес – ізогамія. Мешкають у невеликих прісних водоймах як епіфіти ниткових водоростей. Відомі у центральній Європі та Азії.
Порядок Трибонемові – Tribonematales. Представники мають нитчастий, різнонітчастий, ложнотканинний та тканинний тип диференціації талому. Клітинні стінки або з Н-подібними частинами, що перекриваються, або цілісні.
Рід Трибонема– нитки, що не гілкуються (рис. 46, 1). Клітини за формою циліндричні або барилоподібні. Клітинна стінка складається з двох половинок, які знаходять краями одна на одну у середині клітини. Оболонки часто шаруваті. Фрагменти ниток завжди закінчуються порожніми половинками Н-подібних фрагментів оболонки, що мають вигляд вилочки. У клітинах кілька жовтувато-зелених пластид, піреноїдів немає. Розмноження вегетативне (фрагментацією ниток), безстатеве (зооспорами та апланоспорами) та статеве (ізогамія), причому апланоспори утворюються частіше, ніж зооспори. Можуть формувати акінети. Мешкають у прісних водах, де особливо рясно розвиваються в холодну пору року.
Порядок Вошерієві – Vauсheriales. Всі представники мають сифональний талл, оогамний статевий процес та синзооспори.
Рід Вошерія(Рис. 46, 3) має слоевище неклітинної будови; її талом досягає завдовжки кількох сантиметрів, прикріплюється до субстрату за допомогою безбарвного різоїду. У нитках відсутні перегородки, більшість талому зайнята вакуоллю, а, по периферії в цитоплазмі розташовані численні ядра і пластиди. Нитки з апікальним зростанням та рідкісним бічним розгалуженням. Перегородки утворюються при пошкодженні талому та для відділення органів розмноження. Безстатеве розмноження здійснюється апланоспорами, синзооспорами, акінетами. Синзооспори формуються по одній у зооспорангії, що відокремлюється від вегетативних клітин перегородкою на кінці нитки. Зооспори багатоядерні та багатоджгутикові. Статевий процес – оогамія. Зигота покривається товстою оболонкою і після періоду спокою проростає у новий талом.
Види Вошеріїшироко поширені в прісних, солонуватих та морських водах, а також у наземних місцеперебуваннях. Вони зустрічаються всіх континентах, включаючи Антарктиду. Формують трав'яно-зелені або темно-зелені переплутані маси - так звані мати, гладкі, стелиться або подушкоподібні. Водні, напівводні, наземні форми. Мешкають у різноманітних місцеперебуваннях: морях, естуаріях, лиманах, солоних маршах, мангрових чагарниках, струмках, каналах, озерах, ставках, на орних землях і болотах.
Значеннягетероконтних водоростей
З відділу гетероконтних водоростей найбільше значення для природних екосистем і для людини мають бурі водорості.
Бурі водорості – основний джерело органічної речовиниу прибережній зоні морів. Їхня біомаса в морях помірних і приполярних зон може досягати кількох десятків кілограмів на квадратний метр. Зарості бурих водоростей створюють умови для живлення та розмноження безлічі прибережних тварин та інших водоростей. Чарльз Дарвін, який спостерігав біля берегів Південної Америки зарості бурої водорості Макроцистису, писав: «Ці величезні підводні ліси Південної півкулі можу порівняти лише з наземними лісами тропічних областей. І все-таки, якби в якійсь країні знищити ліс, то не думаю, щоб при цьому загинула хоча б приблизно така кількість видів тварин, як із знищенням водоростей».
Зарості бурих водоростей служать місцем харчування,укриття та розмноженнябагатьох тварин. Образно кажучи, бурі водорості надають іншим водним організмам «стіл, дах та ясла».
Широко використовуються бурі водорості та людиною. Вони багаті йодомта іншими мікроелементами. Народи Південно-Східної Азії традиційно використовують їх в їжу, особливо представників порядку ламінарієвих, з яких готують безліч найрізноманітніших страв. Кормове борошно, що готується з бурих водоростей, підвищує продуктивність худоби; при цьому в яйцях та молоці збільшується вміст йоду.
З бурих водоростей отримуютьальгінати- Солі альгінової кислоти. Альгінати дуже широко застосовують у різних галузях промисловості. Це нетоксичні сполуки, що мають колоїдні властивості, тому вони широко використовуються в харчовій та фармацевтичній промисловості. Альгінова кислота та її солі здатні до 200-300-кратного поглинання води, утворюючи гелі, для яких характерна висока кислотостійкість. У харчовій промисловості вони використовуються переважно як емульгатори, стабілізатори, желюючі та вологоутримуючі компоненти. Наприклад, сухий порошковий альгінат натрію використовують у виробництві порошкоподібних та брикетованих розчинних продуктів (кава, чай, сухе молоко, киселі та ін) для їх швидкого розчинення. Водні розчини альгінатів використовують для заморожування м'ясних та рибних продуктів. У світі харчову промисловість йде до 30% всього обсягу одержуваних альгінатів.
У текстильній та целюлозно-паперовій промисловості альгінати використовують для загущення фарб та посилення міцності їх зв'язку з основою. Просочення тканин деякими альгінатами надає їм захисних властивостей: водонепроникність, кислотостійкість і збільшує механічну міцність. Ряд солей альгінових кислот використовують для отримання штучного шовку. За часів Другої світової війни в США та Англії з альгінової кислоти та її солей вироблялася велика кількість маскувальної тканини та мереж для житлових та промислових будівель.
Альгінати застосовують у металургії: у ливарному виробництві вони покращують якість формувальної землі. Солі альгінової кислоти застосовують у виробництві електродів для електрозварювання, що дозволяють отримувати більш якісні зварні шви. Альгінати також використовують при виробництві пластмас, синтетичних волокон, лакофарбових покриттів та будівельних матеріалів, стійких до атмосферних впливів. Їх застосовують при виготовленні високоякісних мастильних матеріалів для машин. У радіоелектроніці альгінати виконують роль сполучного агента для виготовлення високоякісних феритів.
Найбільше застосування знаходить водорозчинний альгінат натрію, здатний до утворення в'язких розчинів. Його широко використовують для стабілізації різноманітних розчинів та суспензій. Додавання невеликої кількості альгінату натрію в харчові продукти – консерви, морозиво – підвищує їхню якість. Він також використовується для виготовлення декоративної косметики, кремів та масок у парфумерній промисловості.
У фармацевтичній промисловості альгінати використовуються для покриття таблеток, пігулок, як компонентні основи для різних мазей і паст, як гелі-носії лікарських препаратів, у виробництві розчинних хірургічних ниток. У медицині альгінат кальцію застосовують як кровоспинний засіб і як сорбент, що виводить радіонукліди (наприклад, стронцій). Щорічне виробництво альгінатів у світі перевищує 20 тисяч тонн.
Інша важлива речовина, що отримується з бурих водоростей, – шестиатомний спирт маніт. Маніт використовують як замінник цукру для хворих на діабет. Крім того, він може бути використаний як плазмозамінник при консервації крові. Його застосовують для виготовлення таблеток у фармацевтичній промисловості. Маніт також використовують у виробництві синтетичних смол, фарб, паперу, вибухових речовин при виробленні шкір.
Фукоїдани,одержувані з бурих водоростей – ефективні антикоагулянти, навіть активніші, ніж гепарин. Перспективним вважається використання їх для отримання протипухлинних препаратів та антивірусних сполук. Адже навіть у найнижчих концентраціях фукоїдани можуть інгібувати прикріплення вірусів до поверхні клітин. Фукоїдани також мають здатність утворювати виключно міцні і в'язкі слизи, що знаходить застосування в отриманні стабільних емульсій та суспензій.
Енергетична криза, що охопила в останні роки багато країн світу, призвела до необхідності пошуку нових нетрадиційних джерел енергії. Так, у США з цією метою вивчається можливість розведення гігантської ламінарієвої водорості Макроцистисз подальшою переробкою її в метан. Підраховано, що з площі 400 км2, зайнятої цією водорістю, можна отримати 620 млн. м3 метану.
Гетероконтні водорості із класів Золотисті, Жовтозелені, Синурові, Рафідофітові та Євстигмові водорості представлені, в основному, мікроскопічними організмами, широко поширені в прісних водоймах усіх кліматичних зон земної кулі, але частіше зустрічаються в помірних широтах. Є серед золотистих водоростей види, що мешкають у морях та солоних озерах, а дуже мало хто – у забруднених водах. Максимуму розвитку золотисті водорості досягають у холодну пору року: у планктоні вони домінують ранньою весною, пізно восени та взимку. У цей час вони відіграють значну роль як продуценти первинної продукції та служать їжею організмам зоопланктону.
Деякі золотаві водорості, наприклад УрогленаАвтогамія – статеве розмноження, при якому зливаються два сестринські гаплоїдні ядра в загальній цитоплазмі. Динобріон,розвиваючись у масовій кількості, здатні викликати цвітіння води. Вони виділяють альдегіди та кетони, які можуть надавати воді неприємного запаху та смаку. Уроглена– токсичні для риб жирні кислоти
Рафідові водорості широко представлені в планктоні прісних водойм переважно з кислим рН, особливо в сфагнових болотах, рідше у великих озерах. У прісних водоймах локальні «цвітіння» може формувати Гоніостомум.Рафідові водорості зустрічаються також у опріснених морських бухтах та калюжах на березі моря, а також у відкритому морі. При масовому розвитку у прибережних морських водах вони викликають токсичне "цвітіння" води. Так, біля узбережжя Канади під час "цвітіння" концентрація клітин рафідової водорості Гетеросигми може досягати 30 млн. на 1 л. Спалахи чисельності рафідових водоростей часто призводять до розвитку «червоних припливів», з якими пов'язують загибель риб. Причиною таких «червоних припливів» можуть бути види пологів Хаттонелла, Олістодіскус, Гетеросигма та Фіброкапса.
Синурові водорості при масовому розвитку в прісних водоймах можуть надавати воді неприємного запаху ( Синура). Феотамнієві водорості зустрічаються у стоячих та повільно поточних прісних водоймах, де поселяються епіфітно на ниткових водоростях.
Євстигмові водорості зустрічаються лише у прісних водоймах чи грунті.
Жовтозелені водорості поширені на всіх континентах, вони мешкають переважно в прісних водах і ґрунті, а також у наземних, солонуватоводних та морських місцеперебуваннях. Жовтозелені водорості населяють чисті та забруднені води, з різним значенням рН: можуть мешкати як у кислих, так і в лужних водах. Вони зустрічаються, головним чином, у чистих прісноводних водоймищах, рідше в морях і солонуватих водах, віддаючи перевагу помірній температурі, частіше розвиваються навесні і восени, хоча є види, що зустрічаються протягом усіх періодів року, у тому числі і взимку. Найчастіше їх можна знайти в скупченнях нитчаток і серед чагарників вищих водних рослин у прибережній зоні річок, ставків, озер та водосховищ.
Переважна більшість жовтозелених – вільноживучі форми, але зустрічаються і внутрішньоклітинні симбіонти – зооксантели у найпростіших клітинах. Цікавий внутрішньоклітинний симбіоз утворюють хлоропласти морського виду Вошеріїз молюском Елізія. Упродовж дев'яти місяців цей молюск здатний у культурі до фотоавтотрофної фіксації вуглекислого газу. Це найтриваліший у часі симбіоз такого типу, коли симбіотична пластида знаходиться у безпосередньому контакті з цитоплазмою тварини. У природі личинки молюска живляться нитками Вошерії. Внаслідок фагоцитозу хлоропласти водорості потрапляють у цитоплазму епітеліальних клітин молюска. В ході цього процесу оболонка хлоропласту стає тришаровою, втрачається одна зовнішня мембрана ХЕС (хлоропластної ендоплазматичної мережі). Подібне явище є добрим доказом того, що в ході еволюції в результаті вторинного симбіогенезу за рахунок втрати мембран могли виникати хлоропласти з трьома мембранами.
Жовтозелені, золотисті та інші гетероконтні водорості є продуцентами кисню та органічних речовин; вони входять до складу харчових кіл. Гетероконтофіти беруть участь у самоочищенні забруднених вод та ґрунтів, утворенні сапропелю, у процесі накопичення органічних речовин у ґрунті, впливаючи на його родючість. Вони використовують як показові організми щодо стану забруднення вод; жовто-зелені водорості входять до складу комплексу мікроорганізмів, що застосовується для очищення стічних вод.
Контрольні питання
Назвіть характерні риси будови бурих водоростей.
Особливості будови таломів бурих водоростей.
Як розмножуються бурі водорості? Що таке моноспори, тетраспори та зооспори, ізогамія, гетерогамія та оогамія?
Які життєві цикли у бурих водоростей? Розмноження фукусових та ламінарієвих водоростей.
Назвіть характерні риси та типових представників порядків бурих водоростей.
У яких місцепроживання зустрічаються бурі водорості? Яке значення їх у природі.
У яких місцепроживання зустрічаються бурі водорості? Яке значення їх у природі.
Назвіть характерні риси будови та типових представників золотистих водоростей.
Які пігменти та типи харчування відомі у золотистих водоростей?
Розмноження та екологія золотистих водоростей.
Розмноження та екологія золотистих водоростей.
Які пігменти та типи харчування відомі у жовтозелених водоростей?
Які пігменти та типи харчування відомі у жовтозелених водоростей?
Назвіть характерні риси будови та типових представників синурових водоростей.
Назвіть характерні риси будови та типових представників синурових водоростей.
Назвіть характерні риси будови та типових представників феотамнієвих водоростей.
Назвіть характерні риси будови та типових представників феотамнієвих водоростей.
Розмноження та екологія феотамнієвих водоростей.
Назвіть загальні риси будови та типових представників рафідофітових водоростей.
Розмноження та екологія рафідофітових водоростей.
Назвіть загальні риси будови та типових представників евстигмових водоростей.
Розмноження та екологія евстигмових водоростей.
Навчальний посібник написаний відповідно до вимог ФГОС ВПО за напрямом «Педагогічна освіта» та доповнює знання студентів з теоретичної частини курсу «Ботаніка» (систематика рослин та грибів). Матеріал посібника може бути використаний студентами як для самостійної роботи, так і для роботи в аудиторії під керівництвом викладача.
* * *
Наведений ознайомлювальний фрагмент книги Ботаніка. Систематика рослин: навчальний посібник (С. К. П'ятуніна, 2013)наданий нашим книжковим партнером-компанією ЛітРес.
Водорості (Algae)
Численна та різноманітна група нижчих таломних рослин, первинним середовищем проживання яких є вода. Водорості об'єднують кілька самостійних і, ймовірно, незалежно еволюціонували відділів. Представники відділів відрізняються за набором пігментів, деталями тонкої структури хроматофорів, за продуктами фотосинтезу, що накопичуються в клітині (запасних речовин), і за будовою джгутикового апарату. Нижчі рослини – одноклітинні, колоніальні чи ценобіальні та багатоклітинні організми. Цінобіями називають колонії, в яких число клітин визначається на ранніх стадіях розвитку та не змінюється до наступної стадії репродукції (відтворення). Зростання цінобію відбувається за рахунок збільшення розмірів клітин, а не їх числа. Розрізняють такі типи морфологічної організації талому:
1. Монадний– клітини, що активно рухаються за допомогою джгутиків.
2. Кокоїдний- Нерухливі клітини.
3. Ризоподіальний (амебоїдний)- Вегетативні клітини не вкриті оболонками і можуть розвивати цитоплазматичні відростки - ризоподії.
4. Пальмеллоїдний,або капсальний,тип організації представлений нерухомими клітинами, зануреними у загальну слиз.
5. Нитчастий- Клітини, з'єднані в нитки, прості або розгалужені.
6. Гетеротрихальний, або різнонітчастий,- Ускладнений варіант нитчастої будови, для якого характерні дві системи ниток: стелиться по субстрату і вертикальні нитки, що відходять від них.
7. Пластинчастий- Таломи у вигляді платівок.
8. Сифональний- Таломи, часто великих розмірів, формально є однією клітиною зазвичай з великим числом ядер.
9. Сифонокладальнаорганізація представлена багатоядерними клітинами, з'єднаними в нитчасті чи іншій форми багатоклітинні таломи. На перших етапах утворення талому має сифональний тип будови.
Водорості можуть розмножуватися трьома способами: вегетативним, безстатевим та статевим. Вегетативне розмноження полягає у відділенні від цілої рослини частини вегетативного талому, що дає початок новому талому. Безстатеве розмноження здійснюється за допомогою спеціалізованих клітин - суперечка,що утворюються в спорангіях. Суперечки бувають рухливими (Зооспори)або нерухомими (Апланоспори).За формою вони можуть бути ідентичними батьківським таломам (Автоспориодноклітинних водоростей) або різко від них відрізнятися (одноклітинні суперечки багатоклітинних водоростей).
Статеве відтворення у водоростей надзвичайно різноманітне. Найбільш прості форми статевого процесу - злиття морфологічно нерозрізнених вегетативних особин - хологаміяі кон'югація.У значної частини водоростей відбувається утворення спеціалізованих статевих клітин. гамет.Розрізняють таку поведінку гемет:
1. Ізогамію -злиття однакових за формою та розмірами гамет.
2. Гетерогамію –обидва типи копулюючих гамет мають джгутики, але жіноча більша і менш рухлива, ніж чоловіча.
3. Оогамію -злиття нерухомої жіночої яйцеклітини та рухомої чоловічої клітини. Копулюють гамети, що виникли на одній особині (гомоталізм) або на різних особинах (гетероталізм). Гетероталізм спостерігається за будь-якої форми статевого процесу. У ізогамних форм гамети при морфологічному тотожності виявляються фізіологічно різними і позначаються умовними знаками "+" та "-". Чоловічі гамети, що мають джгутики, називають сперматозоїдами,не мають джгутиків, але здатні пересуватися за допомогою амебоїдних рухів називають спермаціями.В результаті статевого процесу утворюється диплоїдна клітина – зигота.
Життєвий цикл,або цикл відтворення водоростей,включає вегетативний ріст, безстатеві спороношення, статевий процес, стадії, що покояться. Співвідношення диплоїдної та гаплоїдної фаз у життєвому циклі водоростей неоднакове. В одних випадках проростання зиготи супроводжується редукційним розподілом (мейозом) зиготи (зиготична редукція),при цьому рослини, що розвиваються, виявляються гаплоїдними. У багатьох зелених водоростей зигота – єдина диплоїдна стадія у циклі розвитку, вся вегетативна фаза проходить у них у гаплоїдному стані. Такий життєвий цикл називають моногаплобіонтним.У частини інших водоростей, навпаки, вся вегетативна фаза диплоїдна, гаплоїдна фаза представлена лише гаметами, перед утворенням яких відбувається редукційний поділ ядра (гаметична редукція),життєвий цикл - монодиплобіонтний.У третіх редукційний поділ ядра передує утворенню спор, що розвиваються на диплоїдних таломах. (Спорична редукція).Вони виростають у гаплоїдні рослини, що розмножуються статевим шляхом. (гаметофіти).Після злиття гамет зигота розвивається в диплоїдну рослину, що несе органи безстатевого розмноження ( спорофіти). Таким чином, у цих водоростей має місце чергування поколінь (генерацій): диплоїдного та гаплоїдного. Життєвий цикл - гаплоїдно-диплоїдний.Обидва покоління можуть бути однакові морфологічно ( ізоморфна зміна поколінь)або ж різко відрізнятися на вигляд (Гетероморфна зміна поколінь).
На практичних заняттях вивчають відділи: зелені (Chlorophyta), діатомові (Bacillariophyta або Diatomeae), бурі (Phaeophyta), червоні (Rhodophyta) водорості.
Відділ Зелені водорості (Chlorophyta)
Відділ Зелені водорості – найбільший за кількістю видів (до 20 000 видів) та морфологічно різноманітний відділ водоростей. Тут є і мікроскопічні дрібні, одноклітинні форми (монадні та кокоїдні) і досить складно влаштовані нитчасті, гетеротрихальні, сифональні, сифонокладальні та пластинчасті, що досягають кількох десятків сантиметрів. Широка й область поширення зелених водоростей (вони зустрічаються по всій земній кулі) і широка їхня екологічна амплітуда. Вони живуть у прісних і морських водоймах, дехто живе поза водою. Але при всьому різноманітті зелені водорості мають ряд загальних ознак:
1) пігментний склад: хлорофіл аі в,каротиноїди та ксантофіли;
2) основний запасний продукт вуглеводної природи - крохмаль, відкладається в хроматофорі навколо піреноїду;
3) світлочутливе вічко - стигма, знаходиться в стромі хроматофора;
4) тилакоїди, що несуть пігменти мають тенденцію до утворення стосів;
5) джгутики ізоморфні (однакові за будовою) та ізоконти (рівні за довжиною).
Клас Власне зелені, або Рівножгутикові, водорості (Chlorophyceae, Isocantae)
Для представників цього класу характерне безстатеве розмноження за допомогою нерухомих апланоспор або рухомих зооспор з двома – чотирма, рідше багатьма ізоконтними та ізоморфними джгутиками. Статеві процеси – хологамія або копуляція гамет – ізогамія, гетерогамія, оогамія. Зигота зазвичай проходить стан спокою і проростає при настанні сприятливих умов, причому її диплоїдне ядро відразу ділиться редукційно. Відповідно до ступенів морфологічного диференціювання талому клас ділиться на порядки.
Порядок Вольвоксові (Volvacales)
Порядок включає одноклітинні, колоніальні та ценобіальні водорості забезпечені джгутиками, тобто монадної організації.
Рід Хламідомонаду (Chlamydomonas)
Хламидомонада – великий рід, що поєднує близько 500 видів, поширений у природі. Його види можна зустріти в дрібних водоймах, калюжах, канавах, що добре прогріваються. При масовому розвитку викликає цвітіння води, особливо у забруднених органічними речовинами водоймищах. Таллом хламідомонади одноклітинний, монадної організації, тобто, перебуваючи в активному стані, хламідомонади швидко пересуваються за допомогою двох рівних джгутиків, прикріплених до переднього кінця тіла. Фаза активного руху змінюється станом спокою. Це так звана пальмеллеподібна стадія, коли клітини втрачають джгутики, їх оболонки сильно ослизняються і утворюють агрегацію клітин хламідомонад, занурених у загальний слиз. У такому вигляді клітини хламідомонад розмножуються поділом. Потрапляючи у сприятливі умови існування, хламідомонади знову виробляють джгутики та переходять до активного руху.
Хламідомонада має целюлозно-пектинову клітинну оболонку, чашоподібний хроматофор з одним або декількома піреноїдами, розташованими в нижній частині, та світлочутливим оком (стигмою) – у верхній частині. Ядро знаходиться в поглибленні хроматофора, є пара пульсуючих вакуолей. Безстатеве розмноження зооспорами відбувається у сприятливих умовах проживання. Кожна хламідомонада потенційно може розмножуватися як вегетативно, так і безстатевим шляхом, а також брати участь у статевому процесі. При безстатевому розмноженні протопласт ділиться на 4 чи 8 частин, утворюються зооспори. Статевий процес у більшості видів ізогамний. Гамети утворюються так само, як і зооспори, але у більшій кількості (32 або 64). Зигота добре пристосована до перенесення несприятливих умов. Її проростання супроводжується редукційним поділом. Цикл розвитку хламідомонад – моногаплобіонтний.
Рід Вольвокс (Volvox)
Рід Вольвокс - колоніальні або ценобіальні водорості. Невеликий рід вольвокс мешкає у чистих стоячих водоймах, ставках та невеликих озерах. Це найбільш високоорганізований представник порядку вольвоксових. Він являє собою велику, що досягає 2-3 мм в діаметрі кулю, одягнений тонким шаром слизу (інволюкрумом), під яким по периферії кулі в один шар розташовуються двожгутикові клітини. Число їх коливається від 500 до 60 000. Внутрішня порожнина кулі зайнята рідким слизом. Клітини колонії за будовою подібні до клітин хламідомонади. Але оболонка кожної клітини сильно ослизнена, тому протопласти сусідніх клітин віддалені один від одного та цитоплазматичні відростки пронизують товщу ослизненої оболонки. У місцях зіткнення формуються плазмодесми.
У безстатевому розмноженні беруть участь 8-10 клітин, розташованих у задній, щодо напрямку руху, частини сфери. Це гонідії. Серед інших ці клітини виділяються більшими розмірами. При їх розподілі спочатку формується плоска 16-клітинна пластинка (гонічна стадія), подальший поділ призводить до формування незамкнутої сфери з маленьким незамкненим отвором, спрямованим до зовнішньої поверхні батьківської колонії. Клітини нового організму, що формуються, звернені своїми джгутиками всередину сфери. Нормальна орієнтування клітин (з передніми кінцями, спрямованими назовні), досягається шляхом повного вивертання навиворіт незамкнутої сфери, тільки після цього її отвір замикається. Репродуктивні клітини диференціюються дуже рано, так що можна спостерігати всередині материнського організму не тільки дочірні, але й онукові колонії. Звільняються молоді колонії після руйнації материнської.
Клітини, які служать для статевого розмноження, – оогонии і антеридії. Оогонія темно-зеленого кольору значно більша за інші клітини і позбавлена джгутиків. В оогонії розвивається одна велика яйцеклітина. Антеридії формують пакети сперматозоїдів. Підлоговий процес у вольвоксу оогамний. Зустрічаються обох статей і роздільностатеві види, а також гомо-і гетероталличние клони. Формується зигота, що покоїться, яка проростає молодою дочірньою колонією після редукційного поділу диплоїдних ядер. Цикл розвитку моногаплобіонтний.
Об'єкти: нар. хламідомонада, нар. вольвокс.
Хід роботи
1. Розгляньте хламідомонаду спочатку на малому збільшенні (м. увел.), потім більш детально при великому збільшенні (б. увел.) вивчіть нерухомі особини, поспостерігайте рух хламідомонади.
2. Зробіть два малюнки:
а) зовнішній вигляд хламідомонади. Позначте оболонку, хроматофор;
б) схему будови клітини хламідомонади, використовуючи таблицю. Позначте оболонку, цитоплазму, ядро, хроматофор, вічко (стигму), піреноїд, джгутики, пульсуючі вакуолі.
3. Розгляньте при м. повів. та замалюйте з препарату сферичні цінобії вольвоксу з дочірніми колоніями.
4. Зробіть схематичний малюнок, використовуючи таблицю, відобразивши особливості будови цінобію. Позначте протопласти, цитоплазматичні відростки, плазмодесми, джгутики, оогонії, антеридії.
Порядок Хлорококові (Chlorococcales)
Порядок включає одноклітинні та ценобіальні форми з кокоідною організацією клітини.
Рід Хлорокок (Chlorococcum)
Рід Хлорокок містить 38 видів і зустрічається в різноманітних місцеперебуваннях: у воді, як у планктоні, так і в бентосі; у ґрунті, а також на корі дерев, на старих дерев'яних спорудах. Хлорокок входить до складу лишайників.
Це одноклітинна кокоїдна водорість, нерухома у вегетативному стані. Клітини мають чашоподібний хроматофор з піреноїдом, але відсутні джгутики, очі і пульсуючі вакуолі. Ядро знаходиться у виїмці хроматофора. У старих особин можна спостерігати кілька ядер, клітини вкриті товстою целюлозною оболонкою.
Розмножується хлорокок безстатевим шляхом за допомогою двожгутикових зооспор подовженої форми. Протопласт материнської клітини ділиться та формується від 8 до 32 зооспор, які звільняються після руйнування стінки материнської клітини. Період активного руху нетривалий; поплававши деякий час, зооспори втрачають джгутики, одягаються оболонкою, ростуть, досягаючи розмірів, характерних для того чи іншого виду. Статевий процес ізогамний. Цикл розвитку моногаплобіонтний.
Рід Гідродиціон (Hydrodiction)
Гідродиціон невеликий, але поширений рід. Зустрічається в затоках річок, ставків та інших стоячих водоймах, збагачених азотом. Це макроскопічна цінобіальна водорість, складена з великої кількості (до 20 000) клітин. Старі екземпляри досягають метрової довжини, а клітини у них до півтора сантиметра.
Гідродикціон має вигляд замкнутої мережі, що складається з 5-6-кутових осередків, утворених гігантськими клітинами, з'єднаними своїми кінцями. У клітинах дорослих сіточек – одна гігантська вакуоль, цитоплазма розташовується постінно і містить сітчастий хроматофор з численними піреноїдами та велику кількість дрібних ядер. Оболонка целюлозна. Кожна клітина виконує всі функції організму (живлення та розмноження).
Розмноження водяної сіточки безстатеве і статеве. До безстатевого розмноження приступають клітини, які досягли досить великих розмірів (0,2 мм). У протопласті формується така кількість зооспор, яка характерна для цього виду сіточки. Зооспори не залишають материнську клітину, а деякий час переміщуються усередині клітини.
Потім скидають джгутики і складаються в молоду сіточку, склеюючись один з одним тими місцями, де проходять тяжі мікротрубочок. Клітини молодої сіточки одноядерні, з пластинчастим хроматофором, що несе один піреноїд. Деякий час молода сіточка живе під оболонкою материнської клітини, та її розмір швидко збільшується, клітини витягуються. Зрештою, оболонка материнської клітини руйнується, і сіточка приступає до самостійного життя.
Статеве розмноження ізогамне, гамет формується більше, ніж зооспор, і вони значно менші. Зигота пофарбована гематохромом у цегляно-червоний колір. Після періоду спокою зигота ділиться редукційно та проростає чотирма великими зооспорами. Вони малорухливі, скоро втрачають джгутики і знову покриваються товстою, але вже скульптурною оболонкою, перетворюючись на так званий поліедр.Поліедри, оболонки яких несуть відростки, а вміст багатий жировими включеннями, мабуть, мають значення поширення водоростей. Поліедри можуть благополучно переносити висихання, і є, таким чином, другою стадією, що покоїться, в циклі розвитку водяної сіточки.
Рід Хлорелла (Chlorella)
Хлорелла - дуже поширена водорість. У природі вона зустрічається в планктоні та бентосі різних водойм, на грунті, бере участь у освіті тіла лишайників, а також живе в симбіозі з дрібними тваринами, утворюючи так звані зоохлорели. Це одна з водоростей, що культивуються. Завдяки високій швидкості розмноження, хлорела дає великий вихід біомаси.
Хлорелла – це одноклітинна кокоїдна водорість кулястої форми. У протопласті знаходиться дзвоноподібний, з великим поглибленням, хроматофор. У западині хроматофора можна знайти ядро. Розмножується хлорелла лише безстатевим шляхом – автоспорами. Статевий процес невідомий. Цикл розвитку безстатевий, моногаплобіонтний.
Об'єкти: нар. хлорокок, нар. водяна сіточка, нар. хлорелу.
Хід роботи
1. Розгляньте при б. повів. і замалюйте хлорокок. Позначте товсту оболонку (пристосування до аерофільного способу життя), чашоподібний хроматофор.
2. Розгляньте при м. повів. фрагмент ценобію водяної сіточки і замалюйте, показавши 5-6-вугільні клітини, що утворюють цінобій. Позначте клітину цінобію, хроматофори.
3. Розгляньте при б. повів. і замалюйте клітину цінобію з молодим цінобієм усередині клітини. Позначте оболонку материнської клітини, молодий цінобій.
4. Замалюйте хлорелу при б. повів. Позначити товсту оболонку, хроматофор.
Порядок Улотриксові (Ulothrichales)
Порядок об'єднує водорості, що мають тал у вигляді нерозгалуженої нитки, складеної з одноядерних клітин, рідше тал пластинчастий або трубчастий.
Рід Улотрікс (Ulothrix)
Улотрикс - досить великий рід, зустрічається в прісних і трохи солонуватих водоймах, віддає перевагу чистій проточній воді. У прибережній зоні річок і струмків, особливо на територіях з відповідним кліматом, можна зустріти Ulothrix zonata. Улотрикс бентосна, прикріплена водорість, утворює скупчення на прибережних каменях.
Улотрикс – це багатоклітинна нитчаста негілляста водорість. Усі клітини, крім базальної, що служить для прикріплення, однотипні. Вони вкриті тонкою, іноді ослизненою, целюлозною оболонкою. Хроматофор постенні у вигляді замкнутого або незамкнутого пояска з великою кількістю піреноїдів. Ядро у клітинах одне. Число клітин у нитках невизначене, тому що у верхніх частинах клітини постійно діляться в одній площині. Розмножується улотрикс вегетативно (фрагментацією нитки), безстатевим та статевим шляхом. Безстатеве розмноження здійснюється зооспорами, забезпеченими чотирма ізоконтними та ізоморфними джгутиками. Зооспори, проростаючи, утворюють нитки улотриксу. Зооспорангієм потенційно може стати будь-яка, крім базальної клітини нитки. Улотрикс утворює двожгутикові ізогамети. Після злиття формується планозигота (рухлива клітка з чотирма джгутиками), яка потім втрачає джгутики та диференціюється в одноклітинний своєрідний спорофіт – розширене вкрите товстою оболонкою тіло на тонкій ніжці. У такому стані зигота-спорофіт знаходиться деякий час, і потім після редукційного поділу в ній формуються зооспори числом 4-16. Такий життєвий цикл характерний для Ulothrix zonata. У деяких видів зигота проростає в диплоїдну нитку. Таким чином, цикл розвитку у Ulothrix zonata – з гетероморфною зміною багатоклітинного нитчастого гаплоїдного спорогаметофіту та одноклітинного диплоїдного спорофіту.
Рід Ульва (Ulva)
Рід Ульва відомий під назвою «Морський салат», широко поширений, але віддає перевагу дрібним водам. Ульва – морська водорість, але добре переносить опріснення. Вона живе у гирлах річок, мілководних лиманах, морських болотах; добре переносить, а частково і віддає перевагу водам зі значним органічним забрудненням. Місцеве населення використовує їх у їжу, але промислового значення ульва немає.
Таллом ульви – багатоклітинний, пластинчастий, складається з двох шарів клітин, краю пластинки, гофровані внаслідок більш інтенсивних клітинних поділів у крайових, порівняно із серединними, зонах. В основі платівка звужується в короткий черешок з підошвою, якою вона і прикріплюється до твердого субстрату. Диференціація клітин у таломі ульви невелика. Спеціальних репродуктивних органів немає. Потенційно кожна клітина може стати спорангієм у диплоїдного покоління або гаметангії у гаплоїдного. Деякі клітини мають трубчасті вирости, що спускаються вздовж центральної частини талому.
Безстатеве розмноження здійснюється чотирижгутиковими зооспорами, що утворюються на диплоїдних рослинах після редукційного поділу. Проростають зооспори в однорядну нитку, що не гілкується, але ще до початку поділу у джгутики, що втратила, і зооспори, що осіла на грунт, виявляється поляризація. Верхній кінець – товстіший, а нижній – тонкий і витягнутий, з нього згодом формуються структури прикріплення. Підлоговий процес ізо-або гетерогамий, гамети двожгутикові. Цикл розвитку гаплодіплобіонтний із ізоморфною зміною поколінь.
Об'єкти: нар. улотрикс, нар. ульва.
Хід роботи
1. Розгляньте при б. повів. і замалюйте ділянку нитки улотриксу, звернути увагу на будову хромотофора. Позначте клітини талому: оболонку клітини, хроматофор, піреноїди.
2. Замалюйте зовнішній вигляд талому ульви, використовуючи вологий препарат.
Порядок Хетофорові (Chaetophorales)
Порядок включає багатоклітинні нитчасті форми гетеротрихального типу з диференціюванням талому на горизонтальну, поширену субстратом, і вертикальну систему ниток.
Рід Драпарнальдія (Draparnaldia)
Види цього роду вимогливі до чистоти та аерації водойм і віддають перевагу річкам і струмкам. Вони в масі ростуть на досить значних глибинах (10 м), утворюючи там цілі чагарники. Драпарнальдія бентосна – прикріплена водорість нитчастої гетеротрихальної будови. У неї є довгі (необмеженого росту) нитки з пояскоподібним хроматофором з порізаними краями. Хроматофор у таких гілок невеликий щодо загального обсягу клітини, тому клітини магістральних ниток бліді. Від цих ниток відходять мутовками пучки коротких сильнорозгалужених гілочок обмеженого зростання, це асимілятори. Хроматофори в них постінні, великі; клітини зеленого кольору. Кожна коротка нитка закінчується безбарвним довгим волоском. Органи розмноження розміщуються серед асиміляторів. Безстатеве розмноження за допомогою чотирижгутикових зооспор. Статевий процес – ізогамія або гетерогамія. Цикл розвитку моногаплобіонтний.
Рід Трентеполія (Trentepohlia)
Трентеполія – аерофільна наземна водорість, добре пристосована до нестачі вологи. Вона оселяється на корі дерев, каменях, дерев'яних будівлях. Особливо багато видів цього роду виявлено у вологих тропічних та субтропічних областях, де трентеполія нерідко веде епіфітний спосіб життя. Трентеполію легко розпізнати серед інших наземних водоростей за рахунок гематохрому - маслорозчинного каротиноїду, що надає їй цегляно-жовтий колір. Талл трентеполії нитчастий гетеротрихальний. Нитки, що стелиться по субстрату, складаються з округлих або овальних клітин, покритих товстою шаруватою оболонкою. Вони пов'язані один з одним порами з плазмодесмами, але проте нитки, що стелиться, легко розпадаються на короткі фрагменти або окремі клітини. Ці фрагменти та клітини в сухому стані розпорошуються та переносяться вітром. Таким чином, трентеполія має досить ефективний механізм вегетативного розмноження, що особливо важливо за умов нестачі води. У клітинах знаходиться кілька дископодібних або стрічкових хроматофорів, позбавлених піреноїдів, і велика кількість ядер, особливо у старих клітин. Крім горизонтальної є система вертикальних ниток, що складається з більш подовжених клітин. Як горизонтальні, і вертикальні нитки рясно гілкуються з допомогою поділу верхівкових клітин, на останніх утворюються шаруваті ковпачки.
Безстатеве розмноження відбувається дво- або чотирижгутиковими зооспорами, що утворюються в спеціальних верхівкових клітинах, - спорангіях, що сидять на трубчастих клітинах - ніжках. Спорангії відокремлюються і повністю переносяться повітряними течіями. Зооспори утворюються лише в тому випадку, якщо спорангій опиниться у воді. Тоді багатоядерний вміст дуже швидко, за кілька хвилин, розпадається на одноядерні ділянки, і виробляються зооспори. Гаметангії також відрізняються морфологічно від вегетативних клітин, але розташовуються переважно на нитках, що стелиться. На відміну від спорангіїв, кулясті гаметангії не мають ніжок. Гаметангії також переносяться повітряними течіями. Потрапляючи у воду, вони проростають двожгутиковими ізогаметами. Однак копуляція відбувається рідко, і гамети розвиваються партеногенетично (без запліднення). У разі формування зигот вони після періоду спокою проростають зооспорами. Цикл розвитку моногаплобіонтний.
Об'єкти: нар. драпарнальдія, нар. трентеполія.
Хід роботи
1. Розгляньте при м. повів. і замалюйте ділянку гетеротрихального нитчастого талому драпарнальдії, відзначте особливості будови «стволових» ниток та асиміляторів, покажіть відмінність будови хроматофорів у клітинах осьової нитки та у клітинах бічних гілочок. Позначте осьову нитку, нитки асимілятори.
2. Розгляньте при б. повів. і замалюйте клітину осьової нитки та клітину нитки асимілятора. Позначте оболонку клітини, хроматофор.
3. Розглянути за б. повів. та замалювати ділянку нитки трентеполії. Позначте шарувату оболонку, запасні речовини (у вигляді крапель олії, забарвлених гематохромом), дископодібні хроматофори.
Порядок Кладофорові (Cladophorales)
Порядок включає водорості неклітинної будови, гіллясті гіллясті, поділені поперечними перегородками на нерівноцінні сегменти, кожен з яких містить багато ядер. Поперечні перегородки виникають незалежно від розподілу ядер.
Рід Кладофора (Cladophora)
Кладофора – дуже великий і поширений рід, переважно морських, і почасти прісноводних водоростей. Описано близько 150 видів. Зустрічається на мілководдях у смузі прибою, на скелях, що вдаються в море, в лагунах, ставках, озерах. Молоді рослини прикріплені до ґрунту або різних підводних предметів, але пізніше відриваються і плавають, утворюючи великі скупчення або жорстку тину; а також великі (10-15 см в діаметрі) кулясті скупчення. Таллом сильногіллястий, сифонокладальної структури. Сегменти, утворені поперечними перегородками, є клітинами у цьому сенсі. Формування поперечних перегородок якісно відрізняється від цитокінезу і пов'язані з розподілом ядер. Сегменти неоднакові за розміром і містять різну кількість ядер. Сітчастий хроматофор також формується шляхом дотику та злиття вільних спочатку хлоропластів. Зовнішня оболонка товста, целюлозна, ніколи не ослизняється, тому тина кладофори жорстка і не слизька.
Розмноження безстатеве і статеве. Зооспорангієм чи гаметангієм може стати будь-який сегмент водорості. Чотирьохжгутикові зооспори виходять із спорангія через пору і проростають спочатку в міхурове тіло, позбавлене перегородок (сифональна стадія), пізніше виникають поперечні перегородки і відбувається розгалуження ниткоподібних структур. Статевий процес ізогамний. Зигота проростає у диплоїдну рослину. Більшість видів кладофоры, що у морях, ізоморфна зміна поколінь, у разі зооспори утворюються після редукційного поділу (мейозоо-спори), але в прісноводних видів відзначений монодиплобіонтний цикл, коли редукційний поділ передує утворенню гамет.
Об'єкти: нар. скарбниця.
Хід роботи
Розгляньте при б. повів. і замалюйте ділянку талому кладофори, що гілкується, із зооспорангіями. Позначте клітини талому, одноклітинні зооспорангії.
Клас Кон'югати (Conjugatophyceae)
У представників цього класу особливий тип статевого процесу – кон'югація, джгутикових стадій немає, безстатевого розмноження суперечками немає.
Порядок Зигнемові (Zygnematales)
Порядок Зігнемові - нитчасті негіллясті багатоклітинні водорості, їх облизня ослизняется, і тому вони слизькі на дотик. Зигота проростає одним проростком, три решта ядра, що утворилися в процесі редукційного поділу, відмирають.
Рід Спирогира (Spirogyra)
Рід Спірогіра – один із найбільших і найпоширеніших по всій земній кулі: він знайдений навіть в Антарктиді. Рідкісна канава, калюжа, ставок або озеро позбавлені слизової обмацки, що плаває на поверхні тину. Таллом спірогіри - нитчастий, негіллястий, всі клітини в нитці рівноцінні та однотипні. У клітині є один або кілька постінних, спіральних, стрічковоподібних хроматофорів, з великою кількістю піреноїдів, розташованих уздовж поздовжньої осі. Краї хроматофора нерівні. У клітці є одна або кілька вакуолей із клітинним соком. У першому випадку цитоплазма займає постінний стан. За наявності кількох вакуолей, окрім постенного шару цитоплазми, є цитоплазматичні тяжі та центральний цитоплазматичний мішечок, в якому знаходиться велике, добре видиме без забарвлення, ядро. Внутрішній шар клітинної оболонки целюлозний, зовнішній – пектиновий, забезпечує ослизнення та утворення драглистого покриву, що надає ниткам шовковистість. У спірогір, що мешкають у водоймах з сильною течією, виробляються різноманітні різозиди, що утримують водорість на місці. Спирогира, як і інші кон'югати, не має в циклі розвитку джгутикової стадії і не утворює суперечки. Вона розмножується або фрагментацією ниток вегетативно, або статевим шляхом. При кон'югації формуються численні зиготи, які після періоду спокою проростають однією ниткою. З чотирьох гаплоїдних ядер, що утворюються в процесі редукційного поділу, три дрібні відмирають, і залишається одне велике життєздатне ядро. Усі поживні речовини, накопичені в зиготі, йдуть формування одного проростка. Зиготи добре пристосовані до перенесення несприятливих умов. Вони вкриті товстою тришаровою скульптурованою оболонкою. Структура оболонки зигот є важливою таксономічною ознакою.
Об'єкти: нар. спірогіра.
Хід роботи
Розгляньте при м. повів. та замалюйте зовнішній вигляд багатоклітинного талому спірогіри. Позначте клітину талому.
1. Розгляньте при б. повів. та замалюйте клітину спірогіри. Позначте оболонку клітини, хроматофор, піреноїди,
ядро, тяжі цитоплазми.
2. Поспостерігайте та замалюйте різні стадії кон'югаційного процесу.
Позначте стадії: утворення кон'югаційних відростків, стиснення та перетікання протопласту, утворення зиготи.
Порядок Десмідієві (Desmidiales)
Десмідієві – одноклітинні організми або ниткоподібні колонії. Клітина складається з двох рівних половинок - напівклітин. При вегетативному розмноженні кожна напівклітина добудовує другу половинку.
Рід Клостеріум (Closterium)
Клостеріум - це прісноводна бентосна водорість, що вимагає хорошого освітлення, вона мешкає в невеликих водоймах, ставках, тихих затоках річок і в обростання підводних предметів. При масовому розвитку формують слизові оболонки. Клостеріум любить чисту воду, але переносить органічні забруднення, а часом зустрічається і в стічних водах. Це одноклітинна форма, її серпоподібне тіло і двох симетричних половин – напівклітин. Ядро розташоване у центрі, в цитоплазматичному мішечку. Зовнішньої перетяжки, властивої іншим десмідієвим водоростям, клостеріум немає, але внутрішня структура відповідає особливостям представників цього порядку. Клостеріум має два однакові осьові хроматофори, своєрідної будови. Від центрального стрижня радіально відходять кілька пластин так, що на поперечному зрізі хроматофор має вигляд багатопроменевої зірки. Великі піреноїди розташовані вздовж стрижня або розкидані безладно біля основи. Основа хроматофора, звернене до центру клітини, - широке, по кінцях клітини хроматофор конічно звужується. На полюсах клітини знаходяться дві невеликі вакуолі з клітинним соком, в який занурені маленькі кристали гіпсу, що знаходяться в постійному русі (броунівський рух), і особливі слизові тільця. Тришарова оболонка клостеріуму пронизана численними конічними порами. Особливо великі пори розташовуються кінцях клітин. Ці пори виділяють слиз, завдяки якому водорость повільно пересувається. У той час як один кінець тіла прикріплюється до субстрату, інший здійснює коливальні рухи. Потім водорість прикріплюється до субстрату іншим кінцем. Так перекидаючись, клостеріум пересувається у напрямку джерела світла.
Розмноження клостеріуму вегетативне. Клітини діляться поперечно. Кожна дочірня клітина отримує половину материнської з одним хроматофором. Друга половинка, тобто напівклітина, добудовується наново. Спочатку молода напівклітина немає хлоропласту, і лише потім хроматофор старої напівклітини ділиться, і його половина перетворюється на нову напівклітину. Таким чином, кожна особина клостеріуму складається з двох різновікових половинок: однієї старішої та іншої молодшої. Статевий процес – кон'югація двох особин, занурених у загальний слиз. Зигота покрита товстою шаруватою оболонкою і добре пристосована до перенесення несприятливих умов. Усю зиму зиготи перебувають у стані спокою, і довгий час ядра залишаються такими, що не злилися. З зиготи формується два молоді клостеріуми, які отримують по два гаплоїдних мейотичних ядра. Одне, маленьке, незабаром дегенерує. Друге – стає ядром нової особини. Цикл розвитку моногаплобіонтний.
Об'єкти: нар. клостеріум.
Хід роботи
1. Розгляньте клостеріум при м. повів. і спостерігайте його рух.
2. Розгляньте клітину клостеріуму при б. повів. та замалюйте її. Позначте оболонку клітини, ядро, хроматофори, піреноїди, термінальні вакуолі із кристалами гіпсу.
Запитання та завдання
1. Перерахуйте основні типи морфологічної організації зелених водоростей. Охарактеризуйте кожен із типів. Наведіть приклади.
2. Які ознаки притаманні представникам відділу зелених водоростей?
3. На які класи підрозділяють відділ зелені водорості? У чому відмінність представників кожного класу?
4. Що таке колонії та цінобії у зелених водоростей? Наведіть приклади колоніальних та ценобіальних зелених водоростей. У чому відмінності колоніальних та багатоклітинних організмів?
5. Як відбувається вегетативне розмноження зелених водоростей? Наведіть приклади.
6. Як відбувається безстатеве розмноження у зелених водоростей? Наведіть приклади.
7. Які статеві процеси характерні для зелених водоростей? Наведіть приклади.
8. Замалюйте схематично життєвий цикл хламідомонди, улотриксу, ульви, спірогіри. Підпишіть на схемі, яке покоління є спорофітом, а яке гаметофітом, набір хромосом (гаплоїдний або диплоїдний) для кожного покоління та клітин службовців для розмноження, тип редукційного поділу та статевого процесу. Вкажіть характерний для цих водоростей тип життєвого циклу.
9. Заповніть таблицю:
Порівняльна характеристика класів та порядків
10. Охарактеризуйте роль зелених водоростей у житті водойм. Наведіть приклади зелених водоростей, які ведуть наземне існування.
Відділ Діатомові водорості (Bacillariophyta, Diatomeae)
Великий відділ одноклітинних та колоніальних організмів, що поєднують понад 10 000 видів. Діатомові клітини покриті кремнеземовим панциром, що складається з двох половинок, що одягаються один на одного, як кришка на коробку. Більша половинка – епітека та менша – гіпотека. Кожна половинка складається з стулки (денця) та спаяного з нею пояскового кільця (пояску). Причому поясок епітеки накладається на поясок гіпотеки. Під панциром знаходиться пектинова оболонка. Діатомові відносяться до групи бурофарбованих водоростей, для яких характерна наявність хлорофілів аАвтогамія – статеве розмноження, при якому зливаються два сестринські гаплоїдні ядра в загальній цитоплазмі. с,маскуються жовтим пігментом фукоксантином. Запасний продукт вуглеводної природи – хризоламінарин. Монадні клітини – сперматозоїди з одним перистим джгутиком. Розмножуються діатомові вегетативно шляхом поздовжнього поділу клітин з добудовуванням однієї стулки – гіпотеки. Статеві процеси – кон'югація та оогамія. Після статевого процесу формується зигота, що має здатність до зростання (ауксоспора). Діатомові водорості живуть у диплоїдному стані і лише їх гамети гаплоїдні.
Клас Центричні (Centrophyceae)
Клас Центричні поєднує водорості з радіально симетричним панциром та відсутністю шовно-вузликового апарату. Усі центричні водорості нерухомі. Статевий процес – оогамія.
Рід Мелозіра (Melosira)
Мелозіра - це нитчаста колоніальна водорість, що складається з однотипних циліндричних клітин, з'єднаних між собою зіткненням дрібних шипиків, розташованих на округлих поверхнях стулок. Панцирь мелозіри має широкі пояски, тому найчастіше водорість розглядається зі своєї бічної поверхні. У клітинах є кілька лопатевих хроматофорів, розташованих постенно, центр клітини зайнятий великою вакуоллю з клітинним соком.
Мелозір не має шовно-вузлової структури і тому нерухома.
Розмножується розподілом та статевим шляхом. Статевий процес – оогамія. В одних клітинах після редукційного поділу утворюється одна яйцеклітина, в інших – чотири сперматозоїди з одним джгутиком. Зигота покрита тонкою, добре розтяжною пектиновою оболонкою. Інтенсивно зростаючі зиготи називаються ауксоспорами. Так як у статевому процесі беруть участь подрібнені після багаторазових поділів клітини, ауксоспора відновлює їх вихідний обсяг. Досягши певних розмірів, ауксоспора виробляє свій панцир. Цикл розвитку мелозіри монодиплобіонтний.
Об'єкти: нар. мелозіра.
Хід роботи
1. Розгляньте ділянку колонії мелозіри при б. повів. та замалюйте водорость з боку пояска та з боку стулки. Позначте клітину колонії, епітеку, гіпотеку.
2. Знайдіть та замалюйте ауксоспору.
Клас Пінатні (Pennatophyceae)
Клас Пенатні включає водорості з двосторонньою симетрією панцира, що мають шовно-вузликовий апарат, мають здатність пересуватися. Статевий процес – кон'югація.
Рід Піннулярія (Pinnularia)
Рід Піннулярія включає понад 150 видів. Мешкає у прісних, бідних вапном, водоймах. Веде бентосний спосіб життя на дні або в обростання підводних предметів. Піннулярія, як і інші діатомові водорості, має значення як кормова база дрібних тварин і є початковою ланкою харчових ланцюгів у водних екосистемах. Це одноклітинна водорість, що має шовно-вузличну структуру і внаслідок цього рухлива. Серед інших одноклітинних діатомових водоростей піннулярія має великі розміри і тому зручна для вивчення. З пояска панцир має прямокутні контури, а стулки – від лінійних до еліптичних. Кінці стулок здебільшого закруглені, але можуть бути відтягнуті та головчасті. У центрі і на кінцях стулки добре помітні вузлики і два s-подібних щілинних отвори (шов), що йдуть від периферичних вузликів до центрального. По краях стулки добре помітний, особливо на порожніх панцирах, чіткий малюнок із паралельних перегородок – септ,не досягають шва. Клітини одноядерні піннулярії з двома пластинчастими хроматофорами, із загнутими краями. Широким плоским боком хроматофор звернений у бік пояска, а краями виходить на бік стулки. Живі активні клітини піннулярії пофарбовані в жовтувато-бурий колір, оскільки фукоксантин маскує зелені пігменти, але в клітинах, що відмирають, фукоксантин вимивається, і хроматофор стає зеленим. У клітинах є дві вакуолі, розділені центральним цитоплазматичним містком. У ньому міститься ядро. Піннулярія запасає волютин, видимий у світловий мікроскоп як тьмяно блискучі кулясті тільця, і краплі олії. Під панциром клітина одягнена пектинової оболонкою, що ослизняється. Піннулярія, що має шовно-вузловий апарат, активно пересувається, повзаючи субстратом.
Розмножується піннулярія розподілом, що проходить паралельно стулкам. Кожна дочірня стулка отримує одну материнську стулку, друга ж стулка, що добудовується, завжди - гіпотека. Внаслідок такої особливості при кожному розподілі одна дочірня клітина завжди трохи дрібніша за материнську, і в популяції можна зустріти особини різного розміру. Підлоговий процес у піннулярії не виявлено, ауксоспор не утворюється. Можна припустити, що великі клітини діляться частіше, ніж дрібні, а найменші не діляться. Життєвий цикл – монодиплобіонтний.
Об'єкт: нар. піннулярія.
Хід роботи
1. Розгляньте піннулярію при б. повів. Замалюйте клітинку з боку стулки. Позначте оболонку клітини, шов, вузлики, септи, хроматофор.
2. Замалюйте клітинку з боку пояска. Позначте епітеку, гіпотеку.
Запитання та завдання
1. Які рівні та типи морфологічної організації характерні для представників діатомових водоростей?
2. Які особливості будови клітин діатомових водоростей?
3. Яку будову має панцир у діатомових водоростей?
4. Які принципи класифікації діатомових водоростей?
5. Які способи розмноження характерні для діатомових водоростей?
6. Як здійснюються статеві процеси у діатомових водоростей? Що таке ауксоспор?
7. Де мешкають діатомові водорості? Які риси пристосування до планктонного та бентосного способу життя мають діатомові водорості? Наведіть приклади планктонних та бентосних діатомових водоростей.
8. Замалюйте схематично життєвий цикл піннулярії та мелозіри. Підпишіть на схемі, яке покоління є спорофітом, а яке гаметофітом, набір хромосом (гаплоїдний або диплоїдний) для кожного покоління і клітин для розмноження, тип редукційного поділу і статевого процесу. Вкажіть характерний для цих водоростей тип життєвого циклу.
Відділ Бурі водорості (Phaeophyta)
Основними пігментами бурих водоростей є хлорофілі аі с,каротиноїди та ксантофіли, у тому числі і фукоксантин, що маскує зелені пігменти і надає водоростям характерного бурого забарвлення. Запасні продукти - ламінарин (речовина вуглеводної природи); маніт – сахароспирт та у невеликій кількості жири. Феопласти пластинчасті або частіше численні дископодібні (зернистий хроматофор), розташовані в перинуклеарному просторі під зовнішньою мембраною ядра, що охоплює кожен феопласт, формує ендоплазматичну феопластну мережу. Піреноїди виступають над поверхнею у вигляді нирки. У клітинах є одне ядро, великі вакуолі з клітинним соком і дрібні вакуолі, що містять дубильні речовини і називаються фізодами.
Оболонка складається з двох шарів: зовнішнього ослизняющегося, що містить альгінову кислоту, і внутрішнього, утвореного особливим різновидом целюлози - альгулозою. Монадні клітини з двома гетероконтними та гетероморфними джгутиками розташовуються збоку клітини. Передній – довгий перистий, покритий мастигонемами, задній – короткий гладкий. Діапазон морфологічних структур великий: від нитчастих гетеротрихальних – до диференційованих пластинчастих форм тканин. У всіх бурих водоростей, за винятком представників порядку фукусові, у яких відсутнє безстатеве розмноження і є диплобіонтами, спостерігається зміна генерацій: в одних – ізоморфна, в інших – гетероморфна. Ці різні типи життєвого циклу покладено основою сучасного поділу відділу бурих водоростей втричі класу.
За небагатьма винятками бурі водорості – морські, особливо багато представлені в холодних водах північної та південної півкулі.
Клас Ізогенератні (Isogeneratophyceae)
Клас Ізогенератний включає водорості з ізоморфним чергуванням поколінь або з гетероморфним, але з домінуванням гаметофіту.
Порядок Ектокарпові (Ectocarpales)включає гетеротріхальні форми.
Рід Ектокарпус (Ectocarpus)
Ектокарпус – широко поширена морська, бентосна водорость, що зустрічається у всіх широтах у прибережній смузі морів та океанів. Водорість поселяється на камінні та інших підводних предметах, у тому числі і на рослинах. Будучи толерантними до різної солоності води, ектокарпуси беруть участь у обростаннях суден. Широка і температурна амплітуда ектокарпусних місць проживання. Він росте як у холодних, так і в теплих морях, активно функціонує як улітку, так і взимку. Ектокарпус – макроскопічна (до 60 см) водорість, нитчаста гетеротрихальна, що має вигляд гіллястих кущиків.
В підставі розташовані горизонтальні ризиди, що стелиться, що прикріплюють водорість до субстрату. Вертикальні гілочки в основі талому, вкриті корою з ризоїдальних ниток, до верхівки стають тоншими і закінчуються довгими безбарвними клітинами. Наростання відбувається за рахунок поділу клітин, розташованих у різних частинах талому. У клітинах ектокарпусу є невеликі вакуолі з клітинним соком, одне ядро в постінній шарі цитоплазми і кілька стрічкоподібних хроматофорів з піреноїдами. У старіючих клітинах хроматофор стає дископодібним.
Розмноження ектокарпусу безстатеве і статеве. Мейозоо-спори, ниркоподібної форми з двома нерівними джгутиками, що прикріплюються до бічної поверхні, утворюються в одноклітинних спорангіях, що сидять на одноклітинній ніжці. Такі спорангії утворюються на спорофітах диплоїдних рослинах. На гаплоїдних рослин закладаються багатогніздні репродуктивні органи. Кожне гніздо продукує одну репродуктивну клітину монадної організації. Найчастіше ці клітини поводяться як гамети і, зливаючись, дають зиготу. Вони однотипні морфологічно, та їх поведінка відрізняється: фізіологічно жіночі гамети менш рухливі і швидко осідають на дно; фізіологічно чоловічі – активніші. З зиготи виростає диплоїдний спорофіт. Описаний вище цикл відповідає дипло-гаплобіонтному типу з ізоморфним чергуванням поколінь, але ектокарпус має відхилення від цього циклу, наприклад, гамети можуть проростати партеногенетично без запліднення, даючи нові гаплоїдні особини. Таким чином, мабуть, досягається можливо повна адаптація ектокарпусу до умов існування та пояснюється його широка екологічна амплітуда.
Об'єкти: нар. ектокарпус (гербарні екземпляри, мікропрепарати).
Хід роботи
1. Розгляньте гербарні зразки ектокарпусу. Замалюйте зовнішній вигляд гетеротрихального талому ектокарпусу.
2. Розгляньте при б. повів. та замалюйте з препарату ділянки талому ектокарпусу з багатокамерними гаметангіями та однокамерними спорангіями. Позначте клітини талому, гаметангії, спорангій.
Клас Гетерогенератні (Heterogeneratophyceae)
Клас гетерогенератних характеризується гетероморфною зміною поколінь з домінуванням спорофіту та мікроскопічними дрібними гаметофітами.
Включає водорості зі складно влаштованим талом, що має тканинну структуру. Наростання здійснюється за рахунок поділу клітин, розташованих у місці переходу листоподібної частини талому в черешок (інтеркалярне зростання).
Рід Ламінарія (Laminaria)
Відомі та широко поширені в північних морях два види ламінарії: Л. цукрова (L.saccharina (L.) Lamour) та Л. пальчаста (L.digitata (Hudz.Lam)), які у верхній субліторалі утворюють цілі плантації та є промисловими водоростями . Ламінарія носить назву «морська капуста» і є цінним харчовим продуктом, що використовується в годуванні тварин, а також виготовлення різних продуктів харчування та лікарських препаратів для людини.
Ламінарія - велика, складно-диференційована бентосна прикріплена водорість, тканинної будови. Її меристематичні клітини здатні ділитися у трьох взаємно перпендикулярних напрямах та утворювати об'ємні структури, у яких всі клітини пов'язані одна з одною плазмодесмами. Таллом ламінарії диференційований на три чітко помітні частини: потужні кігтевидні різозиди, якими вона прикріплюється до субстрату, радіально симетричну частину так званий черешок та сплощену пластинку. Перші дві структури багаторічні, а платівка відмирає і знову наростає завдяки інтеркалярній меристемі, яка знаходиться у верхній частині черешка. Внутрішня організація тіла ламінарії теж досить складна. Черешок з поверхні має щільно зімкнуті клітини, що містять зерна феопластів. У глибших шарах клітини безбарвні та подовжені у поздовжньому напрямку. У центрі клітини розташовані пухко і утворюють серцевину. Черешок поступово наростає в товщину, і на поперечному зрізі добре видно шари, що нагадують річні кільця. В анатомічній будові платівки теж розрізняють дрібноклітинну кору. У центрі проходить «жилка», клітини якої нагадують за своєю будовою ситоподібні клітини вищих рослин.
Розмножуються ламінарії статевим та безстатевим шляхом. Спорангії розташовуються на поверхні пластинки, утворюючи цілі спороносні ділянки або поля, на яких палісадним шаром розташовані мішкоподібні спорангії та подовжені стерильні відростки-парафізи. У спорангіях після редукційного поділу утворюються двожгутикові ниркоподібні мейозооспори, що проростають у гаплоїдні організми. Жіночі та чоловічі гаметофіти відрізняються від спорофіту. Це мікроскопічні ниткоподібні, слаборозвинені рослини (заростки). Вони недовговічні, їхня основна функція – продукція гамет. Статевий процес – оогамія. Зигота без періоду спокою проростає та розвивається у диплоїдний спорофіт. Таким чином, майже вся активна життєдіяльність ламінарії відбувається у диплоїдному стані. Цикл розвитку – диплогаплобіонтний із гетеротрофним чергуванням поколінь, з домінуванням спорофіту.
Кінець ознайомлювального фрагмента.
Загадки для дітей – ребуси з відповідями чи без, можуть бути прості та складні. Кожні ребуси у картинках для дітей мають відповідати розвитку дитини. Тобто, наприклад ребуси для дітей з відповідями на 8 років повинні відповідати знанням, які ваше




